Polyploïdie
Polyploïdie is een mutatie in de aantallen chromosomen bij eukaryote organismen. Deze getalsmatige afwijking van het bestand aan chromosomen door verandering in het aantal chromosomen of delen daarvan heet ook wel ploïdiemutatie of genoommutatie.
|
Bij de gewone kernfasewisseling in de levenscyclus van eukaryote organismen met geslachtelijke voortplanting is er een afwisseling is van een haploïde fase die begint met de producten van een meiose of reductiedeling, en een diploïde fase die begint met een zygote, het versmeltingsproduct van gameten. In de haploïde fase is er een enkele set chromosomen in de celkernen, en in diploïde fase is er een dubbele set chromosomen in de celkernen. Men spreekt echter van een polyploïde eukaryotische cel of organisme wanneer de celkern meer dan twee sets homologe chromosomen bevat. Van elk chromosoom bevinden zich in de celkern vaak meer dan twee exemplaren van de homologe chromosomen. Het aantal sets dat in de cel aanwezig is wordt de ploïdiegraad genoemd. Is dat aantal groter dan twee, dan is er sprake van polyploïdie.
Er zijn twee hoofdcategorieën van polyploïdie: euploïdie en aneuploïdie.[1] Van euploïdie spreekt men als de vermeerdering of vermindering volledige chromosomensets betreft en van aneuploïdie spreekt men als er slechts enkele chromosomen ontbreken of boventallig aanwezig zijn. Het kan gaan om de geslachtschromosomen (heterosomaal) of om de autosomen (de overige chromosomen).
Polyploïdie komt veelvuldig voor bij planten en maar weinig bij dieren.
Euploïdie en aneuploïdie
| directe voorouder diploïde genoom: ZZ |
||||
| ↙ | ↘ | |||
| soortvorming of speciatie (evolutie) | ||||
| ↙ | ↘ | |||
| diploïde soort A genoom: AA |
diploïde soort B genoom: BB | |||
| ↙ | ↓ | ↘ | ↙ | ↓ |
| autopoly- ploïdie |
bastaardering (hybridisatie) |
autopoly- ploïdie | ||
| ↓ | ↓ | ↓ | ↓ | |
| autotetraploïde AAAA |
hybride AB |
autotetraploïde BB | ||
| ↘ | ↓ | ↓ | ↓ | |
| bastaardering | hybridogene polyploïdie |
|||
| ↓ | ↓ | ↓ | ||
| triploïde AAA |
amfidiploïde AABB |
|||
| ↓ | ↓ | ↓ | ||
| diploïdisatie | diploïdisatie | diploïdisatie | ||
| ↓ | ↓ | ↓ | ||
| diploïde soort AA |
diploïde soort AB |
diploïde BB | ||
| schema van boven naar beneden lezen | ||||
Polyploïdie is een gevolg van ploïdiemutaties, waarbij er of te veel chromosomen of te weinig chromosomen aanwezig zijn in de celkern. Deze mutaties ontstaan door een storing in de opbouw van microtubuli tijdens de mitose of de meiose. Dit heeft weer een gevolg voor de juiste verdeling van de chromosomen over de dochtercellen, maar het heeft geen gevolg voor de genen-samenstelling of de structuur van de chromosomen zelf. Bij polyploïdie kan men twee hoofdgroepen onderscheiden: euploïdie en aneuploïdie.
Euploïdie
Bij euploïdie (vermeerdering of vermindering volledige chromosomensets betreft) zijn er op basis van de herkomst van de chromosomensets twee vormen te onderscheiden: auto(poly)ploïdie en allo(poly)ploïdie.
- Bij autoploïdie zijn de chromosomen van slecht één oudersoort afkomstig.
- genoomverdubbeling ((en) Whole genome duplication, WGD) is de vorm van autoploïdie, waarbij in eerste instantie een tetraploïde genoom ontstaat.
- Bij alloploïdie of ook wel hybridogene polyploïdie zijn de chromosomensets van verschillende oudersoorten afkomstig en ontstaat door hybridisatie (bastaardering). Dit komt meer voor dan autoploïdie.
Aneuploïdie
Bij aneuploïdie zijn slechts enkele chromosomen boventallig of ondertallig, bijvoorbeeld geslachtschromosomen (heterosomen). Aneuploïdie is een onvolledige scheiding van de paren van homologe chromosomen; dit kan het gevolg zijn van non-disjunctie tijdens de meiose. Bij het ontbreken van een geheel chromosomenpaar spreekt men van nullisomi, bij monosomie ontbreekt slechts één chromosoom van een bepaald paar, bij trisomie is er één chromosoom extra, bij tetrasomie en pentasomie zijn er twee, respectievelijk drie chromosomen extra bij een bepaald chromosomenpaar.[2]
- Heterosomale (=de geslachtschromosoom betreffende) monosomie (XØ) komt voor als één geslachtschromosoom van het homologe paar ontbreekt. Zo leidt dit bij muizen tot onvruchtbaarheid, en bij de mens in vrouwen tot het syndroom van Turner.
- De autosomale (=geen geslachtschromosoom betreffende) trisomie 21 bij mensen houdt in dat chromosoom 21 in drievoud aanwezig is. Dit leidt tot het syndroom van Down.
- Heterosomale trisomie komt voor bij vruchtbare en geestelijk en lichamelijk volledig ontwikkelde vrouwen (XXX), maar bij mannen (XXY) veroorzaak heterosomale trisomie het syndroom van Klinefelter. Zelfs varianten met XXXY en XXXXY komen voor.
Bij mozaïektriploidie (mozaïcisme) is slechts een deel van het lichaam is triploïd. De triploïdie ontstaat na de bevruchting in een stadium waarin het embryo al meercellig is. Er zijn verschillende karyotypes mogelijk, bijvoorbeeld XXX/XX of XXY/XY. De levensverwachting hangt af van het aandeel aan triploïd weefsel.
Bepalen van de ploïdiegraad
Het aantal chromosomen kan onder het lichtmicroscoop geteld worden. Sinds enige jaren kan ook het gewicht van het DNA in de celkern bepaald worden, waaruit het aantal chromosomen kan worden afgeleid. Het gedrag van de chromosomen in de anafase van de meiose geeft aanwijzingen over de ploïdie: de homologe chromosomen rangschikken zich in paren in het equatorvlak alvorens ze over beide dochtercellen verdeeld worden. Zijn er in plaats van paren groepen van drie (triades) of vier (tetrades), dan is er sprake van een triploïde resp. tetraploïde. Echter, als de chromosoomsets niet van dezelfde soort afkomstig zijn, dan vindt deze hogere groepering niet plaats.
Een juiste interpretatie is alleen mogelijk met kennis van de andere taxa in de verwantschapsgroep, zoals van de andere variëteiten van de soort of de andere soorten in het ondergeslacht of geslacht. Op grond van de chromosomenaantallen bepaalt men het basisaantal: daarin is x = het basisaantal chromosomen binnen de verwantschapsgroep. Bij soorten zonder mutaties in de ploïdiegraad geldt dan 2n = 2x, het diploïde aantal chromosomen is gelijk aan tweemaal het basisaantal.
Voorkomende graden van polypoïdie zijn triploïd (3 sets, anorthoploïde), tetraploïd (4 sets), pentaploïd (5 sets, anorthoploïde), hexaploïd (6 sets), octoploïd (8 sets), decaploïd (10 sets) en dodecaploïd (12 sets). Ploïdietypen volgens het aantal sets chromosomen in de celkernen met voorbeelden zijn:
| polypoïdie | sets | opmerking |
|---|---|---|
| triploïde | 3 sets, anorthoploïde | komen regelmatig voor bij planten (bijv. zaadloze watermeloen), zelden bij amfibieën, algemeen in de stam Tardigrada (beerdiertjes)[3] |
| tetraploïde | 4 sets; 2n=4x | zijn algemeen bij planten (bijv. brede stekelvaren, katoen Gossypium hirsutum), Salmonidae,[4] |
| pentaploïde | 5 sets, anorthoploïde | vindt men alleen bij hybriden |
| hexaploïde | 6 sets; 2n=6x | zijn bijv. brede eikvaren, tarwe, kiwi[5] |
| octoploïde | 8 sets; 2n=8x | bijv. witte steur, dahlias |
| decaploïde | 10 sets; 2n=10x | bijv. bepaalde aardbeien |
| dodecaploïde | 12 sets; 2n=12x | bijv. de plant Celosia argentea of het invasieve Engels slijkgras[6] of de amfibie Xenopus ruwenzoriensis. |
Voorkomen van polyploïdie
Polyploïdie bij dieren
Bij dieren komt polyploïdie (anders dan als genetische afwijking) betrekkelijk weinig voor.
Triploïdie komt bijvoorbeeld (zelden) voor bij amfibieën, maar algemeen in de stam Tardigrada (beerdiertjes).[3]
Bij de mens kan door een genetische fout triploïdie ontstaan. De meeste embryo's sterven al tijdens de zwangerschap en worden dood geboren. Circa een op de 50.000 wordt levend geboren, maar is dan eveneens weinig levensvatbaar en wordt zelden ouder dan enkele maanden. Hoogst zelden wordt de volwassenheid bereikt. De prognose voor kinderen met digynie (of met mozaïektriploïdie) is iets gunstiger.
- Diandrie (type I / hyperandrische triploïdie) - Twee genomen komen van de vader en een van de moeder (XYY). Dit wordt veroorzaakt door bevruchting met twee zaadcellen of door een zaadcel met een diploïde kern. Het embryo wordt gewoonlijk al gedurende de zwangerschap afgestoten.
- Digynie (type II / hypergynische triploïdie) - Er zijn twee vrouwelijke genomen en éen mannelijke (XXY). Dit wordt veroorzaakt door een delingsfout bij de vorming van de eicellen, of door intracytoplasmatische sperma-injectie. Als de kinderen geboren worden overleven ze vaak slechts enige maanden.
Tetraploïdie komt voor bij zalmen en zuigkarpers, octoploïdie bij de witte steur.
Polyploïdie bij hogere planten
Bij planten komt polyploïdie zeer veel voor. Veel soorten zijn in de evolutie ontstaan door polyploïdie. Polyploïden zijn vaak morfologisch herkenbaar aan[7] grotere cellen, en daardoor grotere organen, aan dikkere en relatief kortere, donkerdere, meer behaarde bladeren, aan grotere huidmondjes en epidermiscellen met meer chloroplasten en aan grotere sporen en pollenkorrels.
Bij veel bedektzadigen en varens zijn oude polyploidisaties gevonden, waarmee is aangetoond dat polyploïdisering in evolutionaire zin geen doodlopende weg is, want dan zouden deze lijnen allang zijn uitgestorven. Sommige onderzoekers gaan er zelfs van uit dat alle bedektzadigen het resultaat van polyploïdisering zijn.[8]
Bij anorthoploïdie hebben de soorten en hybriden een oneven aantal sets chromosomen. Deze ondervinden problemen bij de vorming van geslachtscellen omdat de chromosomen niet evenredig over beide haploïde dochtercellen kunnen worden verdeeld door non-disjunctie. De geslachtscellen krijgen een betrekkelijk willekeurig aantal chromosomen toebedeeld en zijn doorgaans niet tot bevruchting in staat. Verdubbeling van alle chromosoomsets maakt van een diploïde hybride een allo-tetraploïde die niet langer lijdt aan non-disjunctie en vaak weer wel vruchtbare nakomelingen kan produceren.
Evolutie en taxonomie
Polyploïdie en evolutie
Gedurende de 20e eeuw bestond er onduidlijkheid of hybridisatie en polyploidisatie moesten worden gezien als belangrijke evolutionaire krachten dan wel als "evolutionaire ruis" zonder betekenis voor de evolutie.
Moleculaire technieken hebben evenwel aangetoond een polyploïde soort vaak meerdere keren wordt gevormd en genetisch bijzonder divers is door de incorporatie van genetische diversiteit van meerdere populaties van hun diploïde voorouders. Bovendien zijn polyploïde genomen zeer dynamisch en genereren veel nieuwe variatie.
In het bijzonder bij planten is soortvorming door hybridisatie en alloploïdie een belangrijk proces, dat aan een groot aantal soorten ten grondslag ligt. Bij een kruising van twee soorten is hybridisering vaak niet voldoende om weer normaal vruchtbare nakomelingen op te leveren, omdat ten gevolge van non-disjunctie van de chromosomen de geslachtscellen niet gevormd kunnen worden. Verdubbeling van alle chromosoomsets maakt van een diploïde bastaard een allo-tetraploïde die niet langer lijdt aan non-disjunctie en vaak weer wel vruchtbare nakomelingen kan produceren. Met name bij planten ligt dit proces aan een groot aantal soorten ten grondslag.
Bij veel bedektzadigen en varens zijn oude polyploidisaties gevonden, waarmee is aangetoond dat polyploïdisering in evolutionaire zin geen doodlopende weg is, want dan zouden deze lijnen allang zijn uitgestorven. Sommige onderzoekers gaan er zelfs van uit dat alle bedektzadigen het resultaat van polyploïdisering zijn.[9]
Polyploïdie en taxonomie
Polyploïdie is een in de taxonomie en de fylogenie gebruikt begrip binnen verwantschapsgroepen om de relaties binnen groepen van verwante taxa te verhelderen. Een soort (of een ander taxon) heet polyploïde als het aantal chromosomen n van de geslachtscellen of van de gametofyt een meervoud is van het grondgetal x. Hierin is x in een volledige, "nauwe verwantschapsgroep" gelijk aan de laagste n-waarde van de set chromosomen van de geslachtscellen of de gametofyt.[10] Bij "nauwe verwantschapsgroep" van soorten gaat het meestal om een serie of sectie, soms om een geslacht.
- x: grondgetal van de chromosomen (= basisaantal), waarvan het aannemelijk is dat dit ook het oorspronkelijke aantal is binnen de verwantschapsgroep.
- n: het haploïde chromosomenaantal van de geslachtscellen of gametofyt (bij planten)
Het somatische (of sporofytische) aantal chromosomen 2n is daarom gelijk aan 2x en zodoende het laagste aantal somatische chromosomen van de betreffende verwantschapsgroep.
In sommige flora's wordt bij de soorten vermeld of het om bastaarden en of het (allo-)polyploïden gaat. Vaak zijn bastaarden steriel en allopolyploïden niet, zodat het verschil onder andere te zien is aan de sporen of het stuifmeel.
Genoomformules
De verschillende sets chromosomen kunnen inzichtelijk en kort omschreven worden met de genoomformule. Dit is een tekenreeks in hoofdletters, met een letter voor elke chromosoomset. Is een alloploïde soort voortgekomen uit de twee diploïde soorten A (met de genoomformule AA) en B (met de genoomformule BB) dan is de genoomformule van de alloploïde AABB.[11]
Een voorbeeld van allopolyploïde hexaploïdie vormt tarwe, dat chromosomensets heeft afkomstig van drie verschillende vooroudersoorten, vaak aangeduid als AABBDD.
- Het genoom van eenkoorn (2n = 14) wordt weergegeven met A en eenkoorn-planten hebben AA.
- Het genoom van de wilde diploïde grassoort (2n = 14) wordt weergegeven met B en planten hebben BB.
- Het genoom van de harde tarwe wordt weergegeven met AB en planten hebben AABB.
- Het genoom van de diploïde Aegilops squarrosa (2n = 14) wordt weergegeven met D en planten hebben DD.
- Het genoom van de gewone tarwe wordt weergegeven met ABD en planten hebben AABBDD.
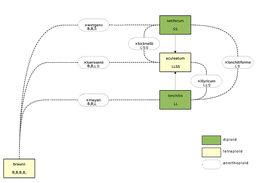
Als ander voorbeeld worden de naaldvarens genomen:
- De zachte naaldvaren Polystichum setiferum is diploïd met de genoomformule SS
- De lansvaren P. lonchitis is diploïd met de genoomformule LL
- de hybride van beide bovenstaande, P. ×lonchitiforme, ontvangt een genoom van beide ouders en heeft dus de genoomformule LS
- De stijve naaldvaren P. aculeatum is tetraploïd, ontstaan door genoomverdubbeling uit bovenstaande hybride en heeft de genoomformule LLSS
- P. braunii is een (segmenteel) autotetraploïde met de genoomformule BBBB
- P. braunii vormt bastaarden met onder meer de stijve naaldvaren: P. ×luerssenii met genoomformule BBLS
Over het algemeen is de plant niet tot normale geslachtelijke voortplanting in staat wanneer de genoomformule niet in twee gelijke delen gesplitst kan worden (LS, BBLS). Wanneer dat wel kan (L+L, S+S, LS+LS, BB+BB), kunnen vruchtbare gameten gevormd worden (met als genoom deze "halve" genoomformules).
Haploïde en diploïde kernfase
Bij de zich geslachtelijk voortplantende organismen is er ook zonder polyploïdie in de levenscyclus steeds een afwisseling van ploïdiegraad. Er is een kernfasewisseling: een haploïde kernfase (aangegeven met n) wisselt af met een diploïde kernfase (aangegeven met 2n). Bij de versmelting van 2 haploïde gameten (zoals een eicel en een zaadcel) ontstaat een diploïde zygote: de bevruchting. Hierbij is het aantal chromosomen in de kern van de zygote het dubbele aantal van dat van de gameten.
Er bestaat een zeer grote variatie in levenscycli. Afhankelijk van de groei en ontwikkeling die plaatsvindt in de haploïde fase en/of in de diploïde fase, zijn de kernfasen kunnen verdeeld over één, twee of soms zelfs drie generaties die elkaar afwisselen.
- Bij dieren is er slechts één diploïde generatie, en alleen de voortplantingscellen zijn haploïde.
- Afwisseling van generaties (generatiewisseling) vindt men onder andere bij zich geslachtelijk voortplantende Embryophyta, zoals mossen, varens en vaatplanten. Er is dan een meer of minder duidelijke haploïde generatie te onderscheiden (de gametofyt), naast een diploïde generatie (de sporofyt).
- Bij sommige groepen van roodwieren zijn er drie generaties te onderscheiden.
Onregelmatigheden in de levenscyclus, met name bij de vorming van de gameten, of bij de bevruchting (bijvoorbeeld bastaardering), kunnen de aanleiding zijn van het ontstaan van polyploïdie.
Zie ook
Bronnen, noten en/of referenties
Bronnen
|
| Mutaties | ||||||
|---|---|---|---|---|---|---|
|
| Het genoom | chromosoom · chromatine · chromatide · histonen · telomeer · centromeer · geslachtschromatine · nucleosoom |
| Chromosomen | karyogram · homologe chromosomen · isochromosoom · autosoom · heterosoom · geslachtschromosoom · X-chromosoom · Y-chromosoom |
| Ploïdie | diploïdisatie · haplogroep · haplotype · haploïdie · diploïdie · dihaploïdie · triploïdie · tetraploïdie · hexaploïdie · octoploïdie · polyploïdie · aneuploïdie · autoploïdie · alloploïdie · euploïdie |
| Genetica | crossing-over · recombinatie · fractionatie · syntenie · mutatie · genoommutatie · puntmutatie · segmentmutatie · basensubstitutie · frameshiftmutatie · deletie · duplicatie · indel · inversie · insertie · translocatie |
| Celdeling | mitose · meiose · karyogamie · spoelfiguur · centrosoom |