Desoxyribonucleïnezuur
Desoxyribonucleïnezuur, afgekort als DNA (Engels: Deoxyribonucleic acid), is een biochemisch macromolecuul dat fungeert als belangrijkste drager van erfelijke informatie in alle bekende organismen, maar ook in virussen (met uitzondering van RNA-virussen). DNA behoort net als RNA tot de nucleïnezuren. Een DNA-molecuul bestaat uit twee lange strengen van nucleotiden, die in de vorm van een dubbele helix met elkaar vervlochten zijn. De twee strengen zijn met elkaar verbonden door waterstofbruggen, die steeds twee tegenover elkaar liggende nucleotiden verbinden. DNA bevat vier verschillende nucleotiden met de nucleobasen adenine, thymine, guanine en cytosine, die afgekort worden met respectievelijk de letters A, T, G en C. De beide strengen zijn complementair doordat de basen alleen als de basenparen A=T en G≡C kunnen voorkomen. A en T worden door 2 waterstofbruggen met elkaar verbonden, G en C door drie waterstofbruggen.
| Portaal Genetica |
De volgorde van nucleotiden in een streng wordt een sequentie genoemd. Doordat er zeer veel sequenties mogelijk zijn, kan de volgorde van nucleotiden unieke erfelijke informatie verschaffen. Zoals met letters woorden, zinnen en boeken gemaakt kunnen worden, zo worden ook de letters van het DNA (de nucleotiden dus) gebruikt om allerlei erfelijke informatie te coderen.
DNA bevindt zich in cellen in de vorm van chromosomen. Chromosomen kunnen miljoenen baseparen bevatten. Door middel van de replicatie wordt het DNA in een chromosoom gekopieerd. De replicatie gaat vooraf aan de celdeling. Zodoende krijgt elke cel een kopie van het DNA, en kan via de voortplanting het DNA doorgegeven worden aan het nageslacht. Op een chromosoom bevinden zich tientallen tot honderden genen. Een gen bestaat uit een of meer DNA-sequenties die coderen voor een of meer eiwitten. Eiwitten vervullen binnen en buiten de cel een zeer grote verscheidenheid aan biologische functies.
In eukaryote organismen bevinden de chromosomen met het DNA zich in de celkern. Daarnaast is er ook DNA aanwezig als mitochondriaal DNA in de mitochondriën in de vorm van circulaire chromosomen, en bij planten daarnaast ook in plastiden (zoals bladgroenkorrels). Prokaryoten (zoals bacteriën en blauwalgen), die geen celkern of andere organellen hebben, bevatten eveneens circulaire chromosomen.
Aan de hand van de genetische code kan de DNA-sequentie van een gen vertaald worden in de aminozuursequentie van een eiwit. Dit proces wordt eiwitexpressie genoemd. Bij de transcriptie wordt het DNA van een gen eerst gekopieerd naar mRNA, en het mRNA wordt vervolgens bij de translatie vertaald naar een eiwit.
Bij de meeste organismen vormen de genen maar een klein gedeelte van de totale hoeveelheid DNA. Veel andere gedeelten van de chromosomen zijn betrokken bij de regulatie van de eiwitexpressie. Van veel van het overige DNA is de functie niet bekend.
Geschiedenis
DNA werd in 1869 ontdekt door de Zwitserse biochemicus Johann Friedrich Miescher (1844-1895). Hij wist de stof te zuiveren uit leukocyten (witte bloedcellen), die hij verkreeg uit pus, afkomstig van ziekenhuisafval.[1] De chemische structuur van DNA bleef nog lange tijd onbekend. In 1909 had Phoebus Levene de theorie opgesteld dat DNA uit vier nucleotiden bestond, en in 1919 publiceerde hij een hypothese voor de wijze waarop de nucleotiden onderling verbonden zijn in een nucleïnezuur.[2]
De natuurkundige Erwin Schrödinger publiceerde in 1944 een invloedrijk boek met de titel Was ist Leben? (Wat is leven?). In dit boek beredeneerde hij dat het erfelijke materiaal moest bestaan uit een aperiodiek kristal: een vaste stof met een regelmatige maar variabele structuur, die de code zou bevatten voor de ontwikkeling van organismen.[3]
Dat het DNA de drager van erfelijke eigenschappen is, werd duidelijk in 1952 door onderzoek van Alfred Hershey en Martha Chase. Zij toonden aan dat een virus, dat bestaat uit DNA (of het verwante RNA) en een eiwitmantel, alleen door middel van DNA een cel kon besmetten. Virussen worden niet gerekend tot de levende organismen. Een virus wordt vermenigvuldigd in een cel.
In 1951 stelde de scheikundige Linus Pauling een model op voor de structuur van DNA. Dit model ging ervan uit dat DNA de structuur had van een helix, maar bevatte op andere punten belangrijke onjuistheden.
In 1952 publiceerde de biochemicus Erwin Chargaff een belangrijke stelregel over de samenstelling van het DNA: dubbelstrengs DNA bevat evenveel adenine als thymine en evenveel cytosine als guanine. Deze regel werd een jaar later verklaard door het concept dat de dubbele helix is opgebouwd uit baseparen van enerzijds adenine en thymine, en anderzijds cytosine en guanine.
 Friedrich Miescher
Friedrich Miescher.jpg) Erwin Schrödinger
Erwin Schrödinger Linus Carl Pauling
Linus Carl Pauling Francis Crick
Francis Crick James Dewey Watson
James Dewey Watson Rosalind Elsie Franklin
Rosalind Elsie Franklin Maurice Wilkins
Maurice Wilkins Frederick Sanger
Frederick Sanger
De correcte chemische structuur van DNA is in 1953 bepaald door het onderzoek van James D. Watson, Francis Crick, Maurice Wilkins en Rosalind Franklin. Franklin deed onderzoek naar de structuur van DNA met behulp van de röntgendiffractietechniek. Een van haar opnamen kwam zonder haar medeweten via Wilkins onder ogen van Watson en Crick. De gegevens van Franklin hielpen Watson en Crick om de structuur van de dubbele helix te bepalen. Wilkins hielp Watson en Crick bij de verificatie van hun model. Watson en Crick publiceerden het artikel over de structuur van het DNA in het tijdschrift Nature op 25 april 1953. In hetzelfde nummer verschenen publicaties van Franklin en Wilkins over de structuur van het DNA. Een maand later publiceerden Watson en Crick op basis van deze structuur een model voor het mechanisme van de replicatie van DNA.[4] Watson, Crick en Wilkins kregen hiervoor in 1962 een Nobelprijs.[5] Franklin was toen al overleden aan kanker.
Francis Crick stelde in 1958 het centrale dogma van de moleculaire biologie op. Dit centrale dogma stelt dat informatie uit genen wél vertaald kan worden naar eiwitten, maar informatie van eiwitten nooit vertaald kan worden naar genen. Het vormde het kader voor de processen van transcriptie en translatie.
In 1975 publiceerde Frederick Sanger een methode voor het bepalen van de volgorde van nucleotiden in DNA. Deze methode, die sequencing wordt genoemd, werd een standaardmethode in de moleculaire biologie. DNA-sequenties werden steeds meer gebruikt om de verwantschap tussen organismen te bepalen. Van steeds meer organismen werd het gehele genoom gesequencet. In 2001 werden de DNA-sequenties van het humane genoom gepubliceerd.
Naamgeving
De afkorting DNA is afkomstig van het Engelse deoxyribonucleic acid, in het Engels zonder s. Dit wordt in het Nederlands vertaald met desoxyribonucleïnezuur. In oudere Nederlandstalige literatuur wordt dit soms als DNZ afgekort. DNA is, zoals de naam al zegt, een nucleïnezuur. Nucleïnezuren zijn ketens van nucleotiden, elk bestaande uit een fosfaatgroep, een ribosegroep en een pyrimidine- of purine-base. Hoewel iedere nucleotide een basische groep bevat, is de bijdrage van de zure fosfaatgroepen groter, zodat DNA toch een zuur genoemd kan worden. Het voorvoegsel 'desoxy-' vertelt dat, in vergelijking met RNA, de ribosegroepen van DNA een zuurstofatoom missen (om precies te zijn, op de zogenaamde 2'-positie van de ribosegroep).
Structuur
| De vier basen:
guanine cytosine adenine thymine |
desoxyribosegroep fosfaatgroep |
Afmetingen
Een DNA-streng is 22 tot 26 ångström breed. Eén nucleotide is 3,3 ångström lang.[6] Een DNA-molecuul kan toch relatief zeer grote afmetingen bereiken. Het langste menselijke chromosoom, chromosoom 1, is bijna 250 miljoen baseparen lang, resulterend in een lengte van ruim 5 cm.[7]
Nucleotiden
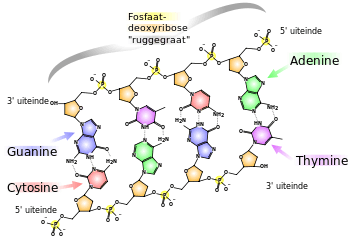
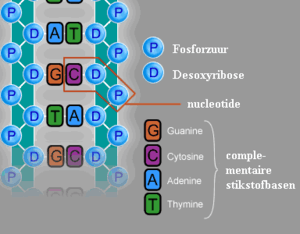
Een DNA-streng is opgebouwd uit nucleotiden. Iedere nucleotide bestaat uit 3 onderdelen: een nucleobase, een desoxyribosegroep (bij RNA: ribose) en 1-3 fosfaatgroepen. In schema:
De basen vormen basenparen en zijn verantwoordelijk voor de erfelijke informatie. De desoxyribose- en de fosfaatgroepen vormen samen de twee "ruggengraten" van het DNA.
Basen en basenparen
In DNA zijn er vier verschillende basen: guanine, cytosine, adenine en thymine, afgekort met respectievelijk G, C, A en T. Een belangrijk onderscheid tussen DNA en RNA is, dat RNA uracil (U) bevat in plaats van thymine.
In het DNA kunnen twee soorten basenparen gevormd worden.
- Guanine en cytosine kunnen samen een basenpaar vormen met drie waterstofbruggen.
- Adenine en thymine kunnen samen een basenpaar vormen met twee waterstofbruggen.
De waterstofbruggen geven een zwakke, niet-covalente binding tussen twee basen. Deze twee baseparen passen als puzzelstukjes in elkaar, de vorming van een basenpaar met een andere base gaat niet of moeilijk.
Verbreking van de basenparen kan relatief makkelijk plaatsvinden door een mechanische kracht of door matige verhitting.[8] Vanwege het aantal waterstofbruggen wordt het GC-basepaar minder gemakkelijk verbroken dan het AT-basepaar. Afhankelijk van de verhouding tussen de GC- en de AT-baseparen wordt dan ook een hogere (veel GC) of lagere (weinig GC) temperatuur gevonden waarbij de twee strengen elkaar loslaten. Deze temperatuur wordt ook wel het smeltpunt van het betreffende DNA genoemd. Voor biologische processen zoals de replicatie en de transcriptie wordt gebruikgemaakt van een helicase om de basenparen te verbreken, en zodoende de twee strengen te scheiden.
Qua chemische structuur worden de basen onderverdeeld in twee soorten:
- Guanine en adenine zijn purinebasen: ze hebben het skelet van purine, dat bestaat uit een 5-ring en een 6-ring.
- Cytosine en thymine zijn pyrimidinebasen: ze hebben het skelet van pyrimidine, dat bestaat uit enkel een 6-ring.
Zowel het purine- als het pyrimidineskelet bevatten meerdere stikstofatomen, waarvan sommige participeren in de vorming van waterstofbruggen. Elk basepaar bestaat uit een purine- en een pyrimidinebase.
 |
 |
De ruggengraten van het DNA
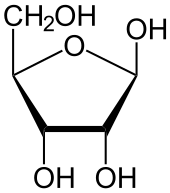
Een DNA-streng kan beschouwd worden als een "ruggengraat" die om en om desoxyribose- en fosfaatgroepen bevat, met de basen als "ribben" die aan de desoxyribosegroepen bevestigd zijn. Door de vlakke structuur van de basen kunnen deze relatief gemakkelijk boven elkaar gestapeld worden.
De desoxyribosegroep is afgeleid van ribose, een suikermolecuul. Op de 2'-positie van de ribose ontbreekt een zuurstofatoom, vandaar desoxyribose. In RNA is er op deze positie een hydroxylgroep, die wel een zuurstofatoom bevat. Deze hydroxylgroep is beschikbaar voor allerlei chemische reacties. Doordat zo'n hydroxylgroep bij DNA ontbreekt, is DNA veel stabieler dan RNA. Dit is een van de oorzaken dat DNA als erfelijk materiaal veel geschikter is dan RNA.
Op de 5'-posities kunnen fosfaatgroepen binden. Op de 3'-posities kunnen de hydroxylgroepen van de suiker binden. Hierdoor kan er een ruggengraat gevormd worden, waarin de desoxyribosegroepen en de fosfaatgroepen om en om zitten.
De twee strengen van het DNA lopen antiparallel: bij de ene streng zit de 5'-positie aan het "begin" en de 3'-positie aan het "eind", bij de andere streng andersom. Het biologische belang van deze oriëntaties is groot: de replicatie en de translatie kunnen alleen in de 5'→3'-richting plaatsvinden.
Een stukje DNA kan nu als volgt worden weergegeven:
| 5' | --- | A | T | G | C | C | G | T | T | A | G | A | C | C | G | T | T | A | G | C | G | G | A | C | C | T | G | A | C | → | 3' |
| 3' | ← | T | A | C | G | G | C | A | A | T | C | T | G | G | C | A | A | T | C | G | C | C | T | G | G | A | C | T | G | --- | 5' |
Dubbele helix
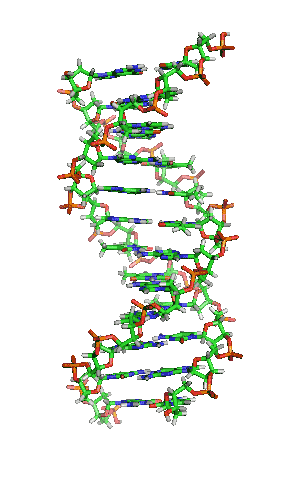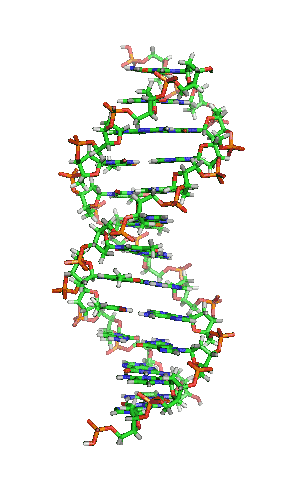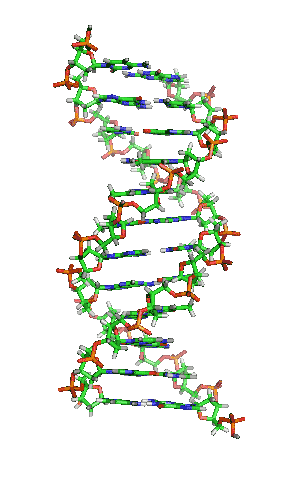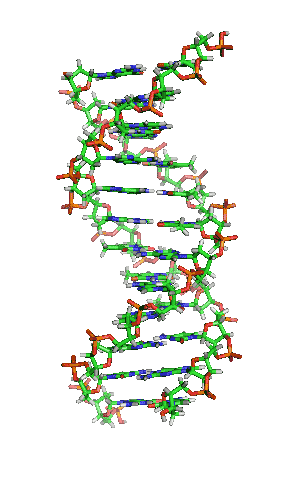
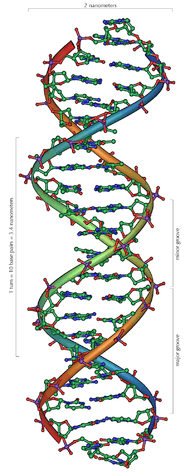
In DNA draaien de twee strengen in een opwaartse spiraal langs elkaar heen. Deze structuur wordt een dubbele helix genoemd. De dubbele helix kan voorkomen in een groot aantal verschillende conformaties. De normale vorm is B-DNA.
B-DNA is rechtsdraaiend. Per draaiing bevat B-DNA ongeveer 10 baseparen. Bij B-DNA hebben de twee groeven tussen de spiralen verschillende afmetingen. Er is sprake van een grote en een kleine groeve (Engels: major en minor groove). De grote groeve heeft een breedte van 22 Å (2,2 nm) en de kleine groeve is 12 Å (1,2 nm) breed.[10]
Een belangrijk effect in de dubbele helix is pi-stacking: de stapeling van basen. Basen hebben een relatief platte vorm door de aanwezigheid van aromatische ringen. Hierdoor kunnen de basen als het ware op elkaar gestapeld worden. Het achterliggende effect is de overlap van de moleculaire orbitalen van de pi-bindingen in de aromatische systemen van aangrenzende basen. De basenvolgorde zou geen invloed op dit effect hebben.[11]
Alternatieve structuren
DNA komt voor in diverse conformatie-isomeren waarvan B-DNA, A-DNA en Z-DNA bekende vormen van de dubbele helix zijn. Daarvan zijn alleen B-DNA en Z-DNA direct als biologisch actief waargenomen. A-DNA schijnt vooral voor te komen in gedehydrateerde monsters van DNA, en is daarom van belang bij kristallografie. Z-DNA komt voor wanneer er veel supercoiling aanwezig is, waardoor er een grote spanning op de DNA-strengen staat. Andere vormen die beschreven zijn, zijn C-DNA, D-DNA, E-DNA, L-DNA, P-DNA en S-DNA.
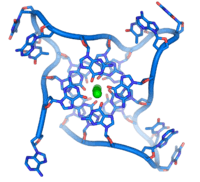
In de grote groeve van B-DNA past een derde streng, waardoor een drievoudige helix gevormd kan worden. Daarbij worden zogenoemde Hoogsteen-baseparen gevormd. Er is maar één geval bekend waarbij een drievoudige helix in vivo voorkomt: in de bacterie E. Coli ontstaat het als een tussenproduct wanneer het recombinatie-enzym RecA actief is.
Aan de uiteinden van chromosomen bevinden zich telomeren, waarin duizenden malen de sequentie TTAGGG wordt herhaald. Dit leidt tot de vorming van een zogenaamde quadruplex van vier DNA-strengen. Hierbij spelen Hoogsteen-baseparen eveneens een rol. Ook op andere plaatsen dan de telomeren kunnen dergelijke quadruplexen voorkomen.
Alternatieve basen
De Nederlandse biochemicus Piet Borst vond een alternatieve base in het DNA: 'Base J'. Deze base is een afgeleide van thymidine.
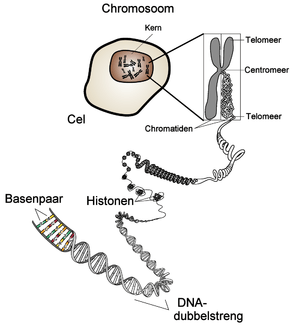
Chromatine
Chromatine is het complex van DNA en eiwitten, die samen de chromosomen vormen. De eiwitten in het chromatine hebben diverse functies: ze helpen het DNA te verpakken in zeer compacte structuren, ze spelen een belangrijke rol bij het regelen van de genexpressie, en ze maken het DNA extra sterk wanneer er tijdens de mitose en de meiose veel spanning op het DNA komt te staan.
Cellen moeten grote hoeveelheden DNA in een klein volume opslaan. Menselijke cellen, bijvoorbeeld, bezitten DNA met een totale lengte van ongeveer 2 meter,[13] dat opgeslagen moet worden in een celkern, die een diameter heeft van 11 tot 22 micrometer.[14] Op verschillende niveaus wordt daartoe het DNA op compacte wijze verpakt.
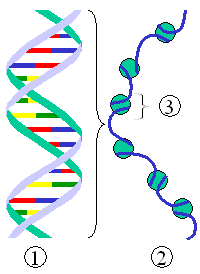
Dat gebeurt in de eerste plaats door de vorming van nucleosomen, die ontstaan wanneer een dubbele helix zich om een aantal histoneiwitten wikkelt. De nucleosomen vormen een soort ketting met kralen, wat in het Engels het beads-on-a-string model wordt genoemd. In ieder nucleosoom is de dubbele helix tweemaal om acht histonen gewikkeld. Tussen de nucleosomen bevindt zich een afstandhouder van koppelings-DNA, een deel van het DNA dat niet gebonden is aan de nucleosomen. De zo gevormde keten van nucleosomen heeft een doorsnede van ongeveer 10 nm (nanometer). Deze keten wordt op nog niet goed begrepen wijze verder verpakt tot een vezel met diameter 30 nm.
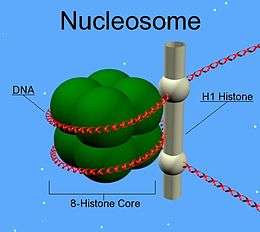
Een nucleosoom bestaat uit een eiwitcomplex van acht histonen en het eromheen gewikkelde DNA. Histonen maken het grootste deel uit van het chromatine. Er zijn vijf soorten histonen, met de namen H1, H2A, H2B, H3 en H4. De histonen H2A, H2B, H3 en H4 komen elk tweemaal voor in een nucleosoom. Deze histonen spelen een belangrijke rol in het regelen van de genexpressie. Acetylering van histonen kan leiden tot activatie van de transcriptie van genen, methylering heeft het tegenovergestelde effect.
Histon H1 maakt geen onderdeel uit van het nucleosoom, het houdt het DNA op z'n plaats en speelt een rol in de vorming van de 30nm-vezel.
De 30 nm-vezel wordt gevormd door een compacte opeenpakking van de nucleosomen in de vorm van een helix waarvan de precieze structuur nog niet bekend is. Ook de 30 nm-vezel wordt weer op compacte wijze gevouwen, waardoor het chromosoom gevormd wordt.
Degradatie van DNA
Nucleïnezuren worden ook in de cel gesplitst of afgebroken. Dit gebeurt met behulp van een enzym, een nuclease. Nucleasen zijn een groep van enzymen, die nucleïnezuren gedeeltelijk of helemaal afbreken. De nucleasen katalyseren de hydrolyse van de chemische binding fosfodi-esterbinding zonder uiteindelijk deel te nemen aan de reactie. Daarom behoren de nucleasen tot de groep katalysatoren.
Functies
Drager van erfelijke informatie

DNA is de drager van erfelijke informatie, opgeslagen in de afzonderlijke genen, die bestaan uit gedeelten van het DNA. Eén gen bevat over het algemeen de instructies voor het synthetiseren van één bepaald eiwit. De eiwitten zijn uiteindelijk bepalend voor de biochemische activiteiten van de cel.
Het aantal genen kan per organisme verschillen. Planten hebben de meeste genen, rond de 50 duizend. De mens en zoogdieren als de muis en de rat hebben ongeveer 25 duizend genen, insecten en wormen tussen de 10 duizend en 20 duizend genen. Alle genen van een organisme worden samen het genoom genoemd.
Bij de celdeling (mitose) wordt het DNA in de oorspronkelijke cel gekopieerd, en elke dochtercel krijgt een kopie van het totale genoom. Bij meiose, een vorm van celdeling waarbij de geslachtscellen (gameten) ontstaan, krijgt elke dochtercel slechts de helft van het DNA in de moedercel. Doordat geslachtscellen wanneer ze versmelten ook hun DNA samenvoegen, heeft het nieuwe organisme dat hieruit ontstaat, weer de normale hoeveelheid DNA. De eigenschappen van het nieuwe organisme zijn op deze wijze van beide ouders afkomstig.
Transcriptie van DNA naar RNA
Het DNA bepaalt de gang van zaken in de cel door het coderen van eiwitten. Dit gaat niet direct, maar met behulp van boodschapper (messenger) RNA ofwel mRNA, dat erg op DNA lijkt. mRNA, dat opgebouwd is uit vrijwel dezelfde bouwstenen (nucleotiden) als DNA, wordt overgeschreven van het DNA (transcriptie). RNA verschilt van DNA doordat het thymine vervangen is door uracil en in plaats van 2-desoxyribose ribose als suiker gebruikt wordt.
Bij de transcriptie bij prokaryoten spelen operons een belangrijke rol. Een operon bestaat uit op het DNA bij elkaar liggende genen die bij elkaar horen voor het tegelijk uitvoeren van één transcriptie van meerdere enzymen voor één celproces. Een voorbeeld is het lac-operon voor de productie van het lactoseverterend enzym (β-galactosidase).
Na de transcriptie verlaten deze mRNA-moleculen de celkern en verplaatsen zich naar andere delen van de cel (het celplasma of naar het endoplasmatisch reticulum). Daar worden de RNA-moleculen vertaald in aminozuren (translatie), die in een lange keten een eiwit vormen. Eiwitten zijn dus ketens van aminozuren, waarvan er in de natuur een twintigtal zijn. Ieder aminozuur in de keten wordt gespecificeerd door een bepaalde volgorde van drie basen (een zogenaamd codon) op een DNA/RNA-streng.
Omdat op elke positie steeds 4 nucleotiden mogelijk zijn, kunnen er 4 × 4 × 4 = 64 combinaties gevormd worden met drie basenparen. Dat zijn er meer dan de voor de aminozuren benodigde 20, dus is de code ontaard: er zijn aminozuren die door meer dan 1 code worden aangeduid. Dit aflezen van de basenvolgorde naar aminozuren wordt gedaan door de ribosomen (grote complexen van RNA en eiwitten). De keten van aminozuren vormt zich op basis van verschillende ladingen en krachten (onder andere H-bruggen, S-bruggen) in die keten tot een eiwit. Vaak wordt een streng meerdere malen afgelezen door een serie van ribosomen, waardoor er in een geringe tijd veel eiwit gemaakt kan worden.
DNA-schade
DNA-schade kan ontstaan ten gevolge van normale biologische processen zoals het metabolisme, maar ook door omgevingsfactoren zoals UV-straling. Naar schatting treden er op moleculair niveau in iedere cel per dag duizenden beschadigingen op aan het DNA. Bij de mens kan dit aantal oplopen tot zo'n 1 miljoen beschadigingen per cel per dag.[15] Veel van deze beschadigingen kunnen zorgen voor schade aan de structuur van het DNA-molecuul. Daardoor kan de transcriptie van een gen beïnvloed worden. Andere beschadigingen leiden tot potentieel gevaarlijke mutaties. Daarom zijn er constant DNA-herstelmechanismen in werking, zodat zo veel mogelijk schade gerepareerd kan worden.
Het grootste deel van de schade aan het DNA betreft de primaire structuur van de dubbele helix. Dat wil zeggen dat er chemische veranderingen optreden aan de basen in het DNA. Ook de dubbele-helixstructuur zelf kan hierdoor aangetast worden, en uiteindelijk ook de structuur van het chromosoom.
DNA-schade kan ook weer ongedaan gemaakt worden door de informatie van de complementaire streng te gebruiken, of door gebruik te maken van de informatie op de zusterchromatide.
Sense en antisense
Als gevolg van het feit dat de beide strengen in tegengestelde richting aan elkaar zitten, en dat de enzymen voor het aflezen van een stuk DNA voorkeur hebben voor een bepaalde richting wordt slechts één streng goed afgelezen voor een bepaald gen. De andere streng kan alleen achterstevoren afgelezen worden, en doordat de nodige sequenties voor transcriptie niet aanwezig zijn op die andere streng zal er geen transcriptie optreden. Moleculair biologen noemen een stuk DNA dat afgelezen wordt of afleesbaar is een antisense en het tegenovergestelde, dus niet afleesbaar, sense (letterlijk vertaald staat sense voor betekenis). Hieruit volgt wat paradoxaal dat de sjabloon (template) voor transcriptie een antisense-streng heeft, hetgeen resulteert in het aflezen van de antisense-DNA-streng en in een sense-RNA-streng.
Onderscheid tussen sense en antisense strengen
Een klein deel van de genen in prokaryoten en een groter gedeelte in plasmiden en virussen maakt het bovenstaande verschil in sense en antisense er niet duidelijker op. Sommige stukken van hun genoom-DNA doen dubbel dienst doordat een eiwit gedecodeerd wordt bij het aflezen van de ene streng van 5' naar 3' en tegelijkertijd een tweede eiwit wordt afgelezen op de andere streng echter ook van 5' naar 3'. Hierdoor kunnen hun genomen in vergelijking met het aantal genen dat ze bevatten zeer compact zijn. Dit bevestigt dat er geen biologisch verschil is tussen de twee strengen, hetgeen nog wordt versterkt doordat elke streng van een dubbele DNA-helix in verschillende gebieden optreedt als sense en antisense.
Enkelstrengs DNA
De meeste virussen hebben geen DNA maar RNA. Er zijn echter ook virussen die wel DNA hebben en bij sommige van deze virussen bestaat het DNA niet uit twee maar uit één streng.
Aanpassing van DNA
DNA kan kunstmatig aangepast worden, maar ook natuurlijk.
Kunstmatige aanpassing van DNA
Tegenwoordig kan DNA kunstmatig aangepast worden door het te extraheren uit een cel, het op te kweken via PCR. Men kan dan proberen het DNA weer in te brengen. Dit wordt vandaag de dag onder andere toegepast bij bacteriën en planten.
DNA kan ook in levende wezens aangepast worden door middel van gentherapie.
Natuurlijke aanpassing van DNA
DNA wordt ook constant door de natuur aangepast, een mogelijkheid is door bijvoorbeeld door recombinatie/crossing-over. Een andere mogelijkheid waardoor het DNA aangepast kan worden is door mutatie, wat ook constant gebeurt in de natuur.
Individuele verschillen in het DNA
Hoewel individuen die tot één soort behoren in het algemeen hetzelfde aantal chromosomen hebben (er zijn uitzonderingen, zoals bij veldbeemdgras) en ook een zeer grote overeenkomst vertonen in hun exacte DNA-volgorde, zijn er tussen individuen van een soort naast de geslachtsverschillen ook altijd enige verschillen aantoonbaar, met name in bepaalde regionen van het chromosoom. Naast polymorfismen (de aan- of afwezigheid van bepaalde kenmerkende stukjes DNA) zijn er variabele regio's (microsatellieten) met lengtevariaties in repeterend DNA, ook wel junk-DNA genoemd, wat het een handig hulpmiddel maakt bij het identificeren van sporen (bijvoorbeeld bloed, sperma, haren) achtergelaten op delictplaatsen. Als men een verdachte heeft, kan men na onderzoek van het spoor en van het DNA van de verdachte nagenoeg met zekerheid zeggen of het spoor van de verdachte afkomstig was of niet.
Ook bij het bepalen van afstamming en bloedverwantschap kunnen deze variabele regio's van het DNA een grote rol spelen. De genetische vingerafdruk, een door mensen gemaakte analyse van de variabele regio's in het DNA, is voor ieder mens uniek mits er voldoende genetische merkers worden gebruikt. Klonen zijn met behulp van de DNA-techniek niet van elkaar te onderscheiden, voor eeneiige tweelingen is een speciale test ontwikkeld om zo ook uitsluitsel te geven.
Bij DNA-onderzoek wordt gewerkt met 11 variabele plekken op een genoom, 1 plek om te bepalen of het DNA-profiel van een man of een vrouw is en de overige 10 om het profiel te maken. Stel dat van elk van deze 10 plekken in het genoom er gemiddeld 10 varianten zijn in een populatie, dan is de kans om twee keer een willekeurig profiel aan te treffen 1 op 10 miljard. De kans dat twee willekeurige individuen hetzelfde DNA-profiel hebben, is daarmee dus verwaarloosbaar klein.
Geografische herkomst
Voor het bepalen van geografische herkomst zijn verschillende methoden beschikbaar, die deels nog in ontwikkeling zijn. De eerste methode kijkt alleen naar het DNA van het Y-chromosoom (het mannelijk geslachtschromosoom). De tweede methode is gebaseerd op het mitochondriaal DNA. De derde en meest recente methode gaat uit van het autosomale DNA. Hierbij kijkt men naar eigenschappen van alle 22 niet geslachtsgebonden chromosomen. Daarbij wordt vooral gelet op specifieke eigenschappen van baseparen, zogenaamde SNP's (single nucleotide polymorphisms).[16] Via deze methode heeft men DNA-verschillen van bewoners van Europese landen in kaart kunnen brengen. Zo heeft de populatiegeneticus Spencer Wells[17] in zijn Genographic Project DNA van volken over de hele wereld verzameld. Dit project is een initiatief van computerbedrijf IBM en de National Geographic Society.[18] Hieraan kan een ieder deelnemen door zijn DNA met een wattenstaafje uit de wang te schrapen en naar het project op te sturen. Met deze methode probeert men menselijke migratiepatronen te reconstrueren op basis van het DNA van mensen die nu leven.[19]
DNA-databank
Er zijn twee soorten DNA-databanken:
- Een bank waarin alle bekende sequenties, stukjes DNA, worden opgeslagen met de daarbij eventueel bekende functie. Hierdoor kunnen onderzoekers een door hen onderzocht stukje DNA vergelijken met alle reeds bekende stukjes. Een overzicht van deze databanken is hier te vinden European Bioinformatics Institute.
- Databanken waarin DNA-profielen zijn opgeborgen om daders van misdrijven te kunnen identificeren, waaronder de DNA-databank voor strafzaken.
DNA in de taxonomie
DNA wordt tegenwoordig veel gebruikt in de systematiek. Het gaat dan in de eerste plaats om chloroplast-DNA bij planten en mitochondriaal DNA bij dieren. Volgens de endosymbiontentheorie is dit in beide gevallen afkomstig van prokaryoot DNA. Een praktische consequentie van deze nieuwe aanpak is het APG III-systeem (2009). Op grond van DNA-sequenties komt men vaak tot andere taxonomische indelingen dan op grond van uiterlijke kenmerken.
Maatschappelijke consequenties van DNA-onderzoek
Het DNA-onderzoek heeft maatschappelijke consequenties, zowel op medisch vlak als op juridisch gebied.
Bij het forensisch onderzoek vormt het DNA-onderzoek een aanvulling op de vingerafdruk. DNA wordt onder andere aangetroffen in bloedsporen en in sperma. Met DNA is het ook mogelijk verwantschap en de etniciteit van de dader vast te stellen, wat met vingerafdrukken niet kan.
Soms wordt aan een groot aantal mensen gevraagd vrijwillig DNA af te staan om te helpen bij het vinden van de dader van een misdrijf. Ook als de dader zelf geen DNA heeft afgestaan, kan men vaststellen of de dader nauw verwant is met iemand die wel DNA heeft afgestaan.
Op die manier kunnen tegenwoordig cold cases alsnog worden opgelost en kunnen rechterlijke dwalingen ontdekt worden, waarbij onschuldigen werden veroordeeld.
Het aantreffen van DNA-materiaal levert niet zonder meer het bewijs van daderschap van degene van wie het DNA is aangetroffen.
Doordat DNA-onderzoek steeds goedkoper wordt is het voor meer mensen mogelijk om verwantschappen vast te stellen, in het bijzonder door het vaststellen van vaderschap.
Synthetisch DNA
Synthetisch DNA (SDNA) wordt gebruikt in DNA-spray dat tijdens een misdrijf gesproeid wordt, en op of in bijvoorbeeld koper om het na koperdiefstal, zelfs na omsmelten, nog te kunnen herkennen.[20]
Dit DNA wordt in een laboratorium opgebouwd uit precies dezelfde bouwstenen A, T, G en C die ook in natuurlijk DNA zijn terug te vinden (zie boven). Een fragment synthetisch DNA bestaat uit tientallen baseparen in een bepaalde combinatie.
Zie ook
| Portaal Genetica |
- Archeogenetica
- Biofotonen
- cDNA
- Chromatine-immunoprecipitatie
- DNA-extractie
- DNA in organismen: eukaryoten, prokarioten, virussen en viroïden.
- DNA-hybridisatie
- DNA-microarray
- DNA-polymerase
- DNA-replicatie (Replicatie (DNA))
- Transposon
- Urinezuur
Bronnen, noten en/of referenties
Bronnen
Externe links |
| Nucleotiden | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
| Zie de categorie DNA van Wikimedia Commons voor mediabestanden over dit onderwerp. |