Celkern
De celkern of nucleus is het door een membraan omsloten deel van een cel waarin het genetisch materiaal (DNA) is opgeslagen. Organismen die een celkern bevatten, worden ingedeeld bij de eukaryoten: de planten, dieren, schimmels en protisten.[kleine-letter 1] Gedurende de celcyclus – het zich herhalende proces van celdeling en groei tot de volgende celdeling – ondergaat de celkern vele functionele en structurele veranderingen.
| Dit artikel is voorgedragen als etalageartikel. Aangemelde gebruikers kunnen gedurende één maand hun stem uitbrengen. |
| Celbiologie |
|---|
| De dierlijke cel |
 |
| Portaal |
De inhoud van de celkern, het kernplasma, wordt omsloten door een dubbel kernmembraan. Om transport van kleine moleculen en kleine eiwitten mogelijk te maken bevat het verder ondoordringbare kernmembraan een aantal kernporiën. Ook grote eiwitten en RNA-moleculen kunnen de kernporiën passeren; daarvoor is echter actief transport nodig met behulp van chemische energie. Binnen de celkern komt het kernlamina voor: een vezelig netwerk dat nodig is voor stevigheid en beweeglijkheid, vergelijkbaar met het cytoskelet.
In de celkern is het meeste DNA van het organisme opgeslagen.[kleine-letter 2] Het DNA is met speciale eiwitten gebundeld tot structuren die aangeduid worden als chromosomen. Het DNA regelt de productie van eiwitten, die maken dat het hele organisme kan functioneren: alle erfelijke eigenschappen worden aangestuurd door informatie uit de celkern. Hierdoor wordt de celkern ook wel gezien als het 'controlecentrum' van de cel. Stoornissen in de organisatie van de celkern zijn van grote invloed op de gezondheid, reden waarom de celkern ook vanuit medisch oogpunt veel is onderzocht.[1]
Hoewel de celkern in het verleden veelal gezien werd als een eenvoudig lichaampje zonder veel structuur, heeft de inhoud van de celkern een uiterst georganiseerde moleculaire architectuur. Onder een elektronenmicroscoop zijn afzonderlijke gebieden van eiwitten, RNA-moleculen en delen van de chromosomen te onderscheiden in een welbepaalde samenhang.[2]
Structuur
Eukaryoten zijn organismen die hun DNA opslaan in een afzonderlijk cellulair compartiment (organel) genaamd celkern of nucleus. Tot de eukaryoten behoren complexe levensvormen: de protisten, schimmels, wieren, planten en dieren, inclusief de mens. In dierlijke cellen is de celkern het grootste organel, dat met behulp van een lichtmicroscoop over het algemeen duidelijk zichtbaar is. Bij zoogdieren bereikt de celkern meestal een grootte van ongeveer 6 micrometer (μm), wat overeenkomt met ongeveer 10 procent van het totale celvolume.[3] De inhoud van de celkern wordt het kernplasma genoemd en is te vergelijken met het cytoplasma van de gehele cel. De dikke grondvloeistof van het kernplasma, het nucleosol, bevat de eiwitten en moleculen die nodig zijn voor het in stand houden en tot expressie brengen van het genetisch materiaal.[4]
Kernmembraan en poriën
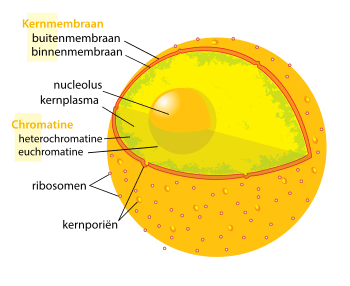
(1) kernmembraan, (2) ringstructuur die de basis vormt van de porie, (3) tussenspaken, (4) korf en
(5) de cytoplasmafilamenten.
(1) kernmembraan, (2) ringstructuur die de basis vormt van de porie, (3) tussenspaken, (4) korf en
(5) de cytoplasmafilamenten.
Het kernmembraan vormt een aaneengesloten barrière rond de celkern en scheidt het genetisch materiaal van het omringende cytoplasma. Het voorkomt dat macromoleculen vrijelijk kunnen diffunderen tussen het kernplasma en het cytoplasma.[5] Het kernmembraan bestaat uit twee parallel aan elkaar afgezette membranen die gescheiden zijn door een tussenruimte van 10 tot 50 nanometer (nm). Het buitenste kernmembraan loopt over in de membraan van het endoplasmatisch reticulum. De ruimte tussen de membranen wordt perinucleaire ruimte genoemd.[5]
Kernporiën, de kanalen die het moleculair verkeer in en uit de celkern verzorgen, zijn gelijkmatig over het kernmembraan verspreid. Ze zijn samengesteld uit verschillende structuureiwitten genaamd nucleoporines. De poriën hebben een molecuulgewicht van ongeveer 125 miljoen dalton en bestaan uit ongeveer vijftig (in gist) tot enkele honderden eiwitten (in gewervelden).[6] Het kanaal waardoorheen moleculen getransporteerd worden is 5–10 nm in diameter. Aan de randen van de opening bevinden zich verschillende regulerende systemen. Via de openingen kunnen kleine, hydrofiele moleculen passief bewegen. Grotere moleculen, zoals nucleïnezuren en eiwitten, kunnen de poriën niet vrijelijk passeren. Deze moleculen worden actief – dat wil zeggen onder verbruik van energie – in en uit de celkern vervoerd.
Bij zoogdieren bevat de celkern ongeveer 3000 tot 4000 kernporiën.[kleine-letter 3] De poriën zijn opgebouwd uit een grote, achtvoudig-symmetrische ringstructuur.[7] Deze ring ligt op de plek waar de binnenste en buitenste membranen fuseren (in elkaar omklappen) en zo een opening vormen. Aan de ring is een korfvormige structuur gehecht die zich uitstrekt naar het kernplasma, en acht dradige filamenten die in het cytoplasma reiken. Beide structuren dienen om passage van transporteiwitten te begeleiden.[6]
Het transport van eiwitten, ribosoomdelen en nucleïnezuren door de kernporiën wordt gecontroleerd door transportfactoren die karyoferinen worden genoemd. Karyoferinen die het verkeer naar de kern verzorgen heten importines; karyoferinen die het verkeer uit de kern verzorgen, heten exportines. Karyoferinen binden aan de te transporteren moleculen in het cytoplasma door herkenning van een signaalsequentie.[8] Na binding vervoeren ze het molecuul via interacties met de reguleringssytemen door de poriën heen.
Nucleair lamina
In dierlijke cellen is de celkern verstevigd door het zogenaamde nucleair lamina, ook wel kernlamina genoemd. Dit is een vezelig netwerk van intermediaire filamenten dat nodig is voor het handhaven van structuur, vorm en beweeglijkheid. Ook speelt het lamina een rol bij het tot expressie brengen van het genetisch materiaal.[1] Bij sessiele organismen zoals schimmels en planten is het kernlamina afwezig.[kleine-letter 4] Het kernlamina staat in verbinding met het cytoskelet via membraaneiwitten aan de buitenkant van de celkern. Het lamina vormt een verankeringspunt voor vele structuren, zowel inwendige (chromatine en chromosomen), als uitwendige (kernporiën).[3]
Het kernlamina is voornamelijk opgebouwd uit het eiwit lamine. Laminen zijn ketens van gevlochten monomeren die gevormd worden in het cytoplasma.[kleine-letter 5] Via poriën gaan de ketens de celkern in, waar ze na samenvoeging met andere eiwitten worden opgenomen in het kernlamina. Ook aan de buitenkant van de kernmembraan binden laminen vast, zoals emerinen en nesprinen. Zij gaan interacties aan met het cytoskelet en zorgen daarmee dat de celkern zich op een vaste plek binnen het cytoplasma positioneert.
Het kernlamina is georganiseerd in een regelmatige structuur die de nucleoplasmatische sluier wordt genoemd. Deze structuur is goed zichtbaar te maken met behulp van fluorescentiemicroscopie.[11] De precieze functie van de sluier is niet geheel duidelijk, maar er zijn aanwijzingen dat deze een rol speelt bij het reguleren van genexpressie.[12] Wanneer de bouwstenen waaruit de sluier bestaat geïnactiveerd worden, leidt dit tot een verminderde interactie met het DNA, waardoor transcriptie van eiwitcoderende genen afneemt.[13]
Het kan voorkomen dat de genen die voor laminen coderen muteren, waardoor deze eiwitten verkeerd worden aangemaakt. Een dergelijk ontwikkelingsdefect van het kernlamina is bij mensen in verband gebracht met enkele zeldzame erfelijke aandoeningen die bekendstaan als laminopathieën. De meest bekende laminopathie is de ziekte progeria, die zich kenmerkt door vroegtijdige verouderingsverschijnselen bij kinderen. De exacte mechanismen die progeria veroorzaken zijn nog grotendeels onduidelijk, maar de meeste onderzoeken suggereren dat de oorzaak ligt bij mutaties in het lamine-eiwit.[1][14]
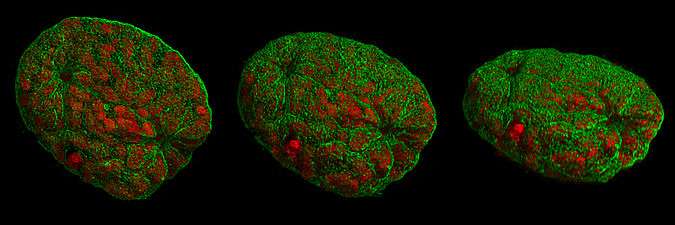
De celkern bevindt zich in een vroeg stadium van deling (profase). De chromosomen (rood) zijn al gecondenseerd voor uiteensplitsing. Het omliggende kernmembraan en inwendige kernlamina (groen) begint te desintegreren. Opgenomen met behulp van 3D-Structured Illumination Microscopy (3D-SIM-microscopy).[15]
Chromosomen
De celkern bevat het grootste deel van het genetisch materiaal van de cel.[kleine-letter 2] Bij eukaryoten is het DNA in de kern rond eiwitten gewonden die histonen worden genoemd. Gedurende het grootste deel van de celcyclus – met name de periode waarin de cel niet deelt – zijn de histonen in elkaar vastgedraaid tot chromatinedraden. Door deze manier van opvouwing is het mogelijk dat iedere celkern een DNA-molecuul van ruwweg twee meter in zich draagt. Wanneer de cel zich gereedmaakt voor de mitose, wordt elk chromosoom gekopieerd. Het gekopieerde DNA trekt zich samen tot dikke structuren, wat goed zichtbaar is onder lichtmicroscopen. Tijdens de celdeling splitsen de chromosomen uiteen in identieke kopieën, zodat twee dochtercellen kunnen ontstaan die allebei beschikken over een identiek genoom.
De verdeling van het chromatine over de celkern is niet uniform door verschillende mate van ontvouwing ten behoeve van DNA-transcriptie. Het chromatine dat minder compact en vrijer in de celkern ligt wordt euchromatine genoemd. Het bevat de genen die veelvuldig door de cel tot expressie worden gebracht en dus voortdurend toegankelijk moeten zijn voor enzymen. Het chromatine dat daarentegen dichter is opgevouwen en zich meer aan de randen van de celkern bevindt wordt heterochromatine genoemd, en bevat het DNA dat weinig of niet door enzymen afgelezen wordt, zoals de sequenties van telomeren of centromeren.[17]
 |  |
Doorsnede van twee dierlijke celkernen.[15] De chromosomen (rood) in profase, het kernmembraan (blauw) en microtubuli (groen) zichtbaar door middel van immunokleuring. De schaalbalk komt overeen met 5 μm. | Ruimtelijke opname van twee dochterkernen.[15] Deze dierlijke dochtercellen zijn in het laatste stadium van de kerndeling: telofase. Het spoelfiguur (bruin), het cytoskelet (groen) en de nog gecondenseerde chromosomen (lichtblauw) zijn zichtbaar in deze SIM-opname. |
Bij een aantal auto-immuunziekten, zoals systemische lupus erythematodes, richten de antilichamen van het lichaam zich tegen bepaalde onderdelen van het chromatine, in het bijzonder tegen de nucleosomen.[18] Deze schadelijke antilichamen staan bekend als antinucleaire antistoffen (ANA) en maken dat het immuunsysteem de eigen celkern aanziet als lichaamsvreemd. ANA’s komen ook voor bij de ziekte multiple sclerose als onderdeel van algemene stoornissen van het immuunsysteem.[19] In het geval van progeria is de rol van antilichamen bij het veroorzaken van auto-immuunsymptomen niet duidelijk.
Nucleolus
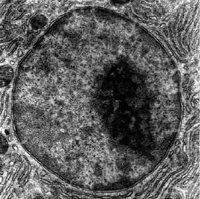
De nucleolus is een grote, amorfe, membraanloze structuur in het kernplasma, die bestaat uit een cluster van DNA, RNA en eiwitten. De nucleolus vormt zich rond specifieke delen van het chromatine, namelijk het DNA dat voor ribosomaal RNA codeert, de zogenaamde rDNA-sequenties. De vorming van dit ribosomaal RNA en de assemblage ervan met r-eiwitten tot ribosomen is de voornaamste functie van de nucleolus.
De grootte van de nucleolus reflecteert zijn activiteit: wanneer veel rRNA wordt gesynthetiseerd in de celkern, zal dit met eiwitten aggregeren tot een samenhangend lichaam. De grootte van de nucleolus is dus sterk afhankelijk van cellulaire activiteit, celgroei en celdifferentiatie. Wanneer het rDNA wordt geïnactiveerd, ontwikkelt de nucleolus zich nauwelijks.[20]
De vorming van ribosomaal RNA gebeurt in de nucleolus met behulp van het enzym RNA-polymerase I. Het rDNA wordt overgeschreven naar een lang pre-rRNA-molecuul, dat vervolgens in een aantal delen gesplitst wordt en zich met ribosomale eiwitten organiseert tot een ribosoomdeel. Deze processen van transcriptie, posttranscriptionele modificatie en assemblage worden ondersteund door kleine RNA-moleculen genaamd snoRNA's.[kleine-letter 6] De ribosoomdelen zijn de grootste structuren die vanuit de celkern doorheen de kernporiën worden getransporteerd. Arriverende in het cytoplasma koppelen de ribosoomdelen aan elkaar en verzorgen dan de eiwitsynthese.
Overige kernlichamen
Hoewel de term kernlichaam in informele zin vaak gebruikt wordt om de nucleolus aan te duiden, zijn er verschillende andere structuren in de celkern ontdekt die als afzonderlijk beschouwd dienen te worden. Belangrijke voorbeelden zijn Cajallichamen, clastosomen en PML-lichamen.[2][21]
Sommige kernlichaampjes kunnen ontstaan als gevolg van bepaalde ziekteprocessen. Dit is bijvoorbeeld het geval bij de spierziekte nemaline myopathie. Deze aandoening wordt veroorzaakt door een mutatie in het eiwit actine. Actine is nodig voor de opbouw van het cytoskelet en is in spiercellen rijkelijk aanwezig. Bij mensen met nemaline myopathie zal het gemuteerde actine in de celkern van spiercellen aggregeren; het klontert samen tot kleine lichaampjes, die met een krachtige microscoop zichtbaar zijn.[22]
Cajallichamen
Over het algemeen bevat de celkern één tot tien compacte Cajallichamen. De diameter van deze lichaampjes kan variëren van 0,2 tot 2,0 μm, afhankelijk van het celtype en de soort. Wanneer ze onder een elektronenmicroscoop worden bekeken lijken ze op een ronde kluwen van draden.[kleine-letter 7] Cajallichamen zijn betrokken bij RNA-processing, verwerking van snoRNA en diverse histon-mRNA-modificaties.[24]
Naast de Cajallichamen komen de vergelijkbare Gemini-lichamen of gems in de celkern voor. Gems komen qua structuur sterk overeen met de Cajallichamen: de twee zijn zelfs met geavanceerde microscopie nauwelijks te onderscheiden.[kleine-letter 8] Het exacte verschil in functie tussen de twee lichaampjes is nog niet opgehelderd.[25] Gems lijken een rol te spelen in het ondersteunen van de vorming van snRNP-moleculen (kleine RNA-complexen die een rol spelen in splicing).[26]
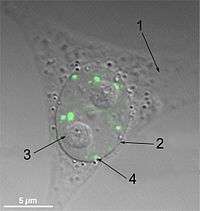
(1) cytoplasma, (2) kernmembraan, (3) nucleolus, (4) paraspeckles.
Clastosomen
Clastosomen zijn kleine, zeldzame kernlichamen (0,2 tot 0,5 μm), die een ringvormig uiterlijk hebben vanwege hun inkapseling.[27] Clastosomen zijn gewoonlijk niet in cellen aanwezig. Ze vormen zich onder sterk proteolytische omstandigheden en verdwijnen weer zodra de proteolyse-activiteit afneemt (bijvoorbeeld na het toedienen van protease-inhibitors). Ook bij osmotische stress kunnen de kernlichaampjes ontstaan.[28] Clastosomen zijn onder andere betrokken bij afbraak van eiwitten.[29]
PML-lichamen
Promyelocytenleukemie-lichamen (PML-lichamen) zijn eiwitrijke structuren die verspreid over het kernplasma voorkomen en 0,1 tot 1,0 μm groot zijn. Ze zijn vernoemd naar hun belangrijkste bestanddeel, het promyelocytenleukemie-eiwit (PML). De lichaampjes kunnen zeer snel veranderen in aantal en chemische samenstelling. Er zijn aanwijzingen dat PLM-lichamen een rol spelen bij het tegengaan van tumorvorming, door schadelijke eiwitten in het kernplasma weg te vangen.[30]
Microscopie
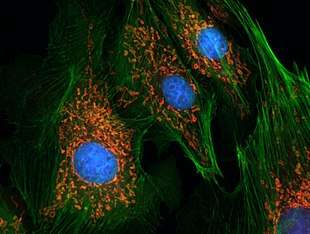
Kennis van de celkern wordt grotendeels verworven aan de hand van microscopisch onderzoek. Er bestaat een breed scala aan microscopische technieken die het mogelijk maakt om cellen, organellen en kleinere structuren in groot detail te bestuderen. Bij fluorescentiemicroscopie worden onderdelen van een cel, zoals het chromatine of het cytoskelet, selectief gekleurd met behulp van een fluorescent molecuul. Omdat bij fluorescentiemicroscopie gewerkt kan worden met levende cellen, kan men begrip krijgen van beweging, interacties en dynamiek van cellulaire processen. Na de ontwikkeling van de elektronenmicroscoop werd de architectuur van de celkern tot op de moleculaire schaal ontrafeld.[31]
- Fluorescentiemicroscopie van cellen waarin de celkern gekleurd is met DAPI
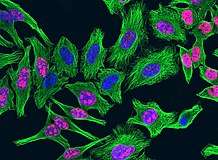 HeLa-cellen. De celkernen fluoresceren blauw. Roze duidt op delingsactiviteit.
HeLa-cellen. De celkernen fluoresceren blauw. Roze duidt op delingsactiviteit.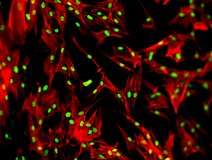 HeLa-cellen. Het cytoskelet is rood gekleurd. Kernen fluoresceren groen.
HeLa-cellen. Het cytoskelet is rood gekleurd. Kernen fluoresceren groen.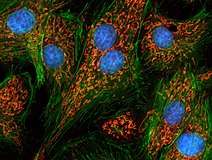 Epitheelcellen van longarteriën. Kernen zijn blauw, mitochondriën oranje.
Epitheelcellen van longarteriën. Kernen zijn blauw, mitochondriën oranje.
Functies
Compartimentalisatie
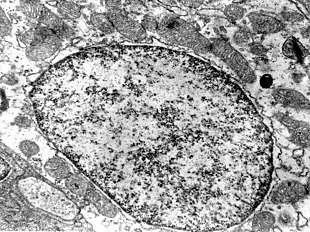
De celkern is in de eerste plaats een afscheidend compartiment: het genetisch materiaal wordt op een gerichte manier van de rest van cel gescheiden, zodat de gevoelige DNA-moleculen afgeschermd worden van de buitenwereld en de processen van transcriptie en DNA-replicatie in een gecontroleerde omgeving kunnen verlopen.[32]
Door de scheiding is er continu communicatieverkeer nodig tussen het cytoplasma en kernplasma. Veel processen in het cytoplasma verlopen onder invloed van enzymen. De informatie voor de aanmaak van enzymen ligt opgeslagen in de celkern, maar de aanmaak zelf (de translatie) vindt plaats buiten de celkern. Wanneer een proces in de cel aan- of uitgeschakeld moet worden, kan een molecuul dat in het proces meespeelt naar de celkern gaan en daar de productie van enzymen reguleren.[kleine-letter 9]
De verschillende RNA-moleculen die in de celkern worden gevormd, ondergaan vaak specifieke modificaties voordat ze naar het cytoplasma worden getransporteerd. Messenger-RNA (mRNA) moet bijvoorbeeld splicing ondergaan, transfer-RNA (tRNA) moet zichzelf lokaal opvouwen tot een klaverbladstructuur en ribosomaal RNA (rRNA) moet zich met eiwitten tot een ribosoom organiseren.[3] Wanneer een onverwerkt RNA-molecuul de celkern verlaat, is deze zelden functioneel.
Instandhouding van DNA
In de celkern vindt replicatie van DNA plaats. Tijdens de replicatie worden de DNA-moleculen die zich in de celkern bevinden volledig gekopieerd, zodat bij de celdeling twee dochtercellen ontstaan die over dezelfde genetische informatie beschikken. Replicatie is een complex proces waarbij veel verschillende eiwitten zijn betrokken. De twee nucleotidestrengen van het DNA worden door het enzym helicase uit elkaar gehaald, waarna in beide richtingen nieuwe complementaire strengen worden gesynthetiseerd door DNA-polymerase. Nadat de replicatie is voltooid kunnen de DNA-moleculen condenseren tot chromosomen en uiteensplitsen voor de celdeling.
In het kernplasma zijn veel enzymen en organische moleculen aanwezig die het DNA voortdurend controleren op beschadigingen. Zulke beschadigingen veranderen de ruimtelijke configuratie van de DNA-helixstructuur en kunnen door DNA-reparatiemoleculen worden opgespoord. Veel van de reparatiemoleculen werken zeer efficiënt, waardoor schade aan DNA sterk wordt beperkt. DNA-reparatie is van essentieel belang voor de integriteit van het genoom. Veel genen die betrokken zijn bij reparatie en bescherming van DNA zijn een bepalende factor voor de levensduur van een individu.[34]
Genexpressie
Genexpressie is het geheel aan moleculaire processen dat ervoor zorgt dat de genetische informatie die in de celkern is opgeslagen, tot uiting komt; oftewel aanleiding geeft tot het ontstaan van waarneembare eigenschappen.[3] De genexpressie wordt meestal aangepast wanneer milieufactoren veranderen (genregulatie). Twee cruciale stappen binnen de genexpressie zijn transcriptie en translatie, die hieronder kort beschreven worden.
Transcriptie is het proces waarbij messenger-RNA (mRNA) wordt gevormd. Langs een stukje DNA (een gen) wordt een complementair RNA-molecuul overgeschreven. Het mRNA is een boodschappermolecuul: het bevat de genetische informatie voor de synthese van eiwitten. In het kernplasma wordt het mRNA-molecuul afgewerkt – het ondergaat splicing, en wordt op de juiste plekken voorzien van extra sequenties – waarna het via de kernporiën de celkern verlaat.
In het cytoplasma wordt het mRNA omsloten door een ribosoom. Hierin vindt de translatie plaats, het proces waarbij de nucleotidevolgorde van het mRNA wordt vertaald naar een volgorde van aminozuren, die door het ribosoom aan elkaar worden gekoppeld. Uit de aminozuurketen ontstaat uiteindelijk een functioneel eiwit. De verzameling van eiwitten die op deze wijze in de cel tot expressie komen, vormt de grondslag van de morfologie en fysiologie van het organisme.
In eukaryoten vinden transcriptie en translatie plaats in afzonderlijke cellulaire compartimenten: transcriptie verloopt binnen de celkern, translatie verloopt in het cytoplasma. Er is sprake van een scheiding in ruimte en in tijd.[35] Bij prokaryoten zijn de twee processen nauw aaneengeschakeld: de translatie van bacterieel mRNA kan al beginnen terwijl de transcriptie nog gaande is. Door de ruimtelijke scheiding van transcriptie en translatie zijn eukaryoten in staat genexpressie op ingewikkelde manieren te reguleren, waardoor er veel meer structuren en functies ontwikkeld kunnen worden dan bij prokaryoten.[35]
Verwerking van mRNA
Een andere belangrijke functie van de celkern is het reguleren van posttranscriptionele modificaties: wijzigingen van het nieuw gevormde messenger-RNA. Nieuw gesynthetiseerde mRNA-moleculen (pre-mRNA) moeten deze modificaties in de celkern ondergaan voordat ze naar het cytoplasma worden vervoerd; mRNA dat zonder deze modificaties in het cytoplasma terechtkomt, wordt direct afgebroken.[36] De drie voornaamste modificaties van pre-mRNA zijn het aanbrengen van een 5'-cap, het toevoegen van een poly(A)-staart en bovenal splicing.
Splicing van mRNA wordt uitgevoerd door een eiwitcomplex dat het spliceosoom wordt genoemd. Tijdens de splicing worden introns – korte regio's in het DNA die niet coderen voor eiwitten – uit het pre-mRNA verwijderd. De resterende exons worden aan elkaar gekoppeld tot een functioneel molecuul. Dit molecuul, het rijpe mRNA, kan door ribosomen worden getransleerd. Doordat exons op verschillende manieren kunnen worden samengevoegd, kunnen verschillende mRNA-moleculen worden gevormd uit een enkel pre-mRNA. Dit proces van alternatieve splicing maakt het mogelijk uit een beperkte hoeveelheid DNA veel verschillende eiwitten te vormen.[6]
Dynamiek en regulatie
Transport
Elk van de kernporiën in het kernmembraan kan per seconde duizenden moleculen gecontroleerd laten passeren.[3] Dit nucleair transport is gebaseerd op een complexe wisselwerking van processen tussen de kern en het cytoplasma. Het transport naar de celkern bestaat voornamelijk uit eiwitten (zoals laminen, DNA- en RNA-polymerase), koolhydraten, signaalmoleculen en lipiden; het uitgaand transport bestaat uit RNA-moleculen en ribosoomdelen. Kleine deeltjes kunnen vrijelijk door de kernporiën heen bewegen door middel van passieve diffusie.[37] Grotere deeltjes passeren de poriën met snelheden die afnemen met het molecuulgewicht: een groot eiwit passeert trager dan een klein eiwit. Voor het transport van grote moleculen is hulp nodig van hulpeiwitten genaamd karyoferinen. Voor de invoer zorgen importines, voor de uitvoer exportines.
De energie die nodig is voor de invoer en uitvoer van stoffen door kernporiën, wordt gehaald uit de concentratiegradiënt van het energierijke molecuul GTP (een stof vergelijkbaar met ATP).[kleine-letter 10] De verbindende factor tussen dit gradiënt en de karyoferinen is het enzym Ran, dat tussen de celkern en het cytoplasma circuleert. Wanneer Ran zich in het kernplasma bevindt, gaat het een binding aan met GTP; in het cytoplasma bindt het met het molecuul GDP. De gevormde Ran-complexen geven het nucleair transport richtingsgevoeligheid: het Ran-GTP-complex zorgt ervoor dat importines hun invoermolecuul loslaten, en dat exportines hun uitvoermolecuul juist binden. Het omgekeerde geldt voor Ran-GDP.[8]
Alle RNA-moleculen die in de celkern worden gevormd, worden naar buiten getransporteerd door gespecialiseerde exporteiwitten. Alleen wanneer alle posttranscriptionele modificaties en verdere afwerkingen aan het RNA zijn voltooid, gaan de exporteiwitten een binding aan met het RNA. Ook deze exporteiwitten maken gebruik van het GTP-gradiënt, maar op basis van een ander mechanisme, aangezien het hier gaat om eenrichtingsverkeer.
Opbouw en afbraak
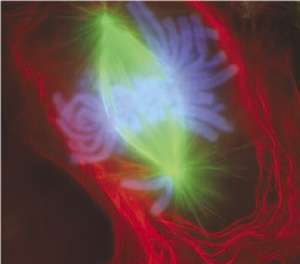
Gedurende één celcyclus vindt volledige opbouw en afbraak van de celkern plaats. Wanneer een cel zich gereedmaakt voor deling, worden de structurele elementen van de celkern – het kernmembraan en het lamina – systematisch ontmanteld. In de meeste eukaryotische cellen markeert het uit elkaar halen van de celkern het einde van de profase (het eerste stadium van de mitose). Bij sommige eencellige eukaryoten, zoals gistcellen, blijft de celkern tijdens de deling intact. De dochterchromosomen bewegen dan naar de tegenovergelegen uiteinden van de celkern, en uiteindelijk splitst deze in tweeën. In hogere eukaryoten worden de chromosomen door een spoelfiguur naar beide uiteinden van de cel getrokken.[kleine-letter 11] Na insnoering en deling van het cytoplasma vormt zich een nieuw kernmembraan rond de chromosomen.
Om er zeker van te zijn dat de microtubuli van het spoelfiguur zich aan de chromosomen kunnen vasthechten, wordt het kernmembraan rond de chromosomen tijdig afgebroken. Als deze intact blijft, komt de mitose niet op gang.[38] Het ontmantelingsproces van het kernmembraan en lamina wordt gereguleerd door eiwitkinases die onder andere de laminen in de celkern voorzien van een fosfaatgroep (fosforylering). De lamine-eiwitten laten van elkaar los waardoor de celkern desintegreert. Tegen het eind van de mitose worden de fosfaatgroepen weer verwijderd, waardoor de structuureiwitten weer bijeenkomen en de celkern zich hervormt.[38]
Kwantiteit
Verreweg de meeste eukaryotische cellen hebben een enkele celkern. Bij gespecialiseerde celtypen kan de celkern echter afwezig zijn, of juist in meervoud voorkomen. Kernloze cellen kunnen ontstaan wanneer de cel door een fout onevenredig deelt, zodat de ene dochtercel een kern mist en de andere twee kernen heeft. Meestal is een afwijkend aantal kernen echter het gevolg van normale processen, zoals bij de kernloze rode bloedcellen. Dwarsgestreepte spierweefsels zijn opgebouwd uit cellen die versmolten zijn, waardoor er meerdere kernen in een individuele cel aanwezig kunnen zijn.[39] Ook bij schimmels en eencelligen is meerkernigheid geen zeldzaam fenomeen.
Kernloze cellen
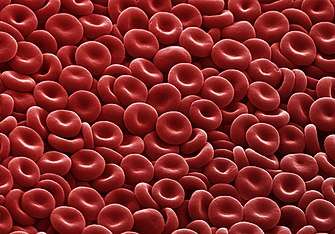
Cellen die geen celkern bevatten zijn uitsluitend opgebouwd uit cytoplasma en zijn daarom niet in staat om dochtercellen te produceren. Vermeldenswaard is de kernloze rode bloedcel of erytrocyt, die als belangrijkste taak heeft zuurstof van de longen naar de lichaamsweefsels te transporteren. Rode bloedcellen ontwikkelen zich in het beenmerg via een proces genaamd erytropoëse, waarbij ze hun kern en organellen verliezen. Tijdens de differentiatie van een erytroblast naar een reticulocyt – de voorlopers van een rode bloedcel – wordt de kern fysiek uit de cel verdreven. Men vermoedt dat dit de cel klein en flexibel genoeg maakt om door haarvaten te kunnen bewegen.[40] Mutagene stoffen kunnen ervoor zorgen dat de bloedcellen in wording te vroeg aan de bloedbaan worden afgegeven. Hierdoor ontstaan er erytrocyten met gereduceerde "microkernen".[41]
In bedektzadigen bestaat een vergelijkbaar proces voor zeefcellen. Zeefcellen vormen lange, aaneengesloten vaten waarlangs water en voedingsstoffen getransporteerd worden. Gedurende de differentiatie van zeefcellen degenereren de celkern, de tonoplast en andere organellen, zodat er in de floëemcellen ruimte ontstaat voor efficiënt watertransport.[42]
Meerkernige cellen
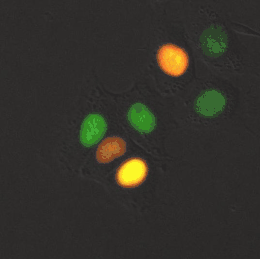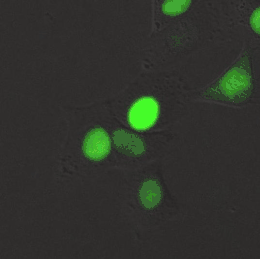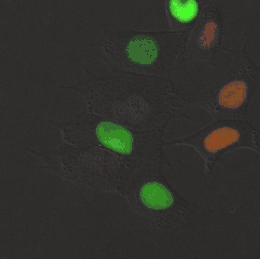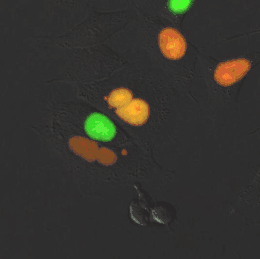
Er bestaan diverse levensvormen die volledig of gedeeltelijk zijn opgebouwd uit meerkernige cellen. Een bekend voorbeeld is Dikarya, een groep hogere schimmels die op een vast stadium in hun levenscyclus tweekernige schimmeldraden vormen.[43] Eencellige eukaryoten (protisten), zoals radiolaria en dinoflagellaten, kunnen ook van nature twee of meer kernen bevatten. Bijzonder aan deze organismen is dat de kernen elk een aparte afkomst hebben: één kern is van de dinoflagellaat en de andere kern van een symbiotische diatomee.[44] Verschillende groenwieren en roodwieren (zoals Griffithsia en Chara) vormen coenocytische cellen waarin meermaals kerndelingen plaatsvinden.[45]
Bij de mens zijn spiercellen van de skeletspieren versmolten tot weefselstructuren die syncytia genoemd worden. In het syncytium kunnen de kernen vrij bewegen. Ze komen voornamelijk voor in het perifere gedeelte van de cellen om ruimte te maken voor de myofibrillen (spiereenheden). Andere meerkernige cellen in het menselijk lichaam zijn de osteoclasten: speciale beencellen die het botweefsel aanleggen en afbreken. Bij ontstekingen en vermoedelijk ook bij de ontwikkeling van tumoren kan het voorkomen dat twee witte bloedcellen (monocyten en macrofagen) fuseren tot een meerkernige reuzencel.[46][47]
Evolutie
Omdat de celkern de definiërende eigenschap is van eukaryoten, is de evolutionaire oorsprong ervan al lange tijd onderwerp van wetenschappelijke discussie. Er zijn verschillende hypothesen voorgelegd die het bestaan van de celkern in evolutionaire termen verklaren, maar geen ervan wordt algemeen geacepteerd.[48] De ontwikkeling van de celkern en andere organellen, zoals het mitochondrion en de chloroplast, zijn cruciale stappen geweest in de evolutie van het leven. Meercelligheid en motiliteit, twee wezenlijke eigenschappen van complex leven, waren een gevolg van een sterke diversificatie en specialisatie op cellulair niveau.
Oorsprong van de celkern
De oorsprong van de celkern kan verklaard worden door middel van het syntrofische model. Deze hypothese gaat uit van een symbiotische associatie tussen bacteriën en archaea, twee oude groepen organismen die van zichzelf geen celkern bevatten. Er wordt verondersteld dat de symbiose tot stand kwam toen ruwweg 2 miljard jaar geleden een voorouderlijk archaeon door een voorouderlijke bacterie werd opgenomen. Door vele generaties samen te leven verloor de archaeon zijn oorspronkelijke stofwisseling. De bacterie groeide en uiteindelijk veranderde de archaeon in een primitieve celkern: een compartiment met RNA en DNA dat een fysiologische en genetische eenheid vormde met de bacterie.[49][50]
Deze ontstaanshypothese van de celkern wordt ondersteund door verschillende argumenten. Ten eerste is het DNA en de genetische machinerie van eukaryoten zeer vergelijkbaar met dat van archaea.[51] Daarnaast zijn er bacteriën bekend die motiliteit vertonen, multicellulaire complexen kunnen vormen en eiwitten bezitten die overeenkomen met die van eukaryoten. Dit deed vermoeden dat eukaryotische cellen een bacteriële oorsprong hebben.[52]
Een tweede hypothese stelt dat eukaryotische cellen ontstaan zijn uit primitieve bacteriën die in structuur overeenkomen met tegenwoordige planctomyceten: bacteriën waarin membraancompartimenten zijn geëvolueerd.[53] In deze bacteriën komt een 'kernmembraan' voor waarin het bacterieel DNA en RNA afgeschermd worden van de rest van de cel. De cellulaire structuur van planctomyceten is een belangrijke aanwijzing voor een niet-symbiotische oorsprong van de celkern.[48]
Endosymbiose
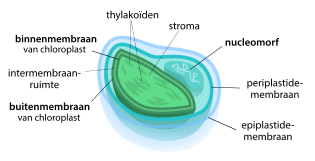
Er zijn sterke aanwijzingen dat mitochondriën en plastiden van oorsprong zelfstandige prokaryoten waren die door een voorouderlijke cel werden opgenomen. In plaats van de opgenomen prokaryoot te verteren, maakte de gastheercel gebruik van de stofwisseling van de prokaryoot, waardoor de twee ingesteld samen gingen leven (endosymbiose). Na verloop van tijd verloor de endosymbiont een groot deel van zijn genetisch materiaal. Veel van zijn genen werden overgedragen naar de kern van de gastheercel,[kleine-letter 12] een verschijnsel dat endosymbiotische genoverdracht wordt genoemd.[54]
Dankzij endosymbiose ontstonden onder meer de eerste eencellige roodwieren en groenwieren. Later in de evolutie ondergingen deze organismen een secundaire endosymbiose: ze werden zelf opgenomen door een andere eukaryotische cel. Een bepaalde groep wieren, de chlorarachniofyten, zou op deze manier ontstaan zijn. De chloroplasten van deze wieren zijn door vier membranen omgeven en bevatten zelfs nog een rudimentaire celkern, de zogenaamde nucleomorf.[55]
Ontdekkingsgeschiedenis

De celkern was een van de eerste celbiologische ontdekkingen. De oudst bekende tekening waarin cellen met een celkern zijn afgebeeld, werd gemaakt door Antoni van Leeuwenhoek (1632–1723) en dateert uit begin 18e eeuw. In een preparaat van rode bloedcellen uit zalmen[kleine-letter 13] constateerde hij een "holte", de celkern.[56]
De celkern werd gedetailleerder beschreven door de Schotse botanicus Robert Brown in 1831. Brown bestudeerde de epidermiscellen van orchideeën onder de microscoop en zag daarbij in iedere cel een ondoorzichtig gebied, dat hij "areola" of "nucleus" noemde.[57] Bij zijn beschrijvingen gaf hij nog geen mogelijke functie van de celkern.
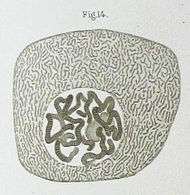
In 1838 kwam Matthias Schleiden op het idee dat de celkern een rol speelt bij het genereren van nieuwe cellen, waarbij hij de naam "cytoblast" (celbouwer) introduceerde. In zijn experimenten beweerde hij te zien dat nieuwe cellen zich rond deze "cytoblasten" verzamelden. Franz Meyen was een sterke tegenstander van deze opvatting. Hij geloofde dat celkernen zeldzaam waren, en verklaarde het vermenigvuldigen van cellen in termen van deling. Het idee dat cellen spontaan uit een "cytoblast" ontstonden was bovendien in tegenspraak met de latere celtheorie van Robert Remak (1852) en Rudolf Virchow (1855), die stelde dat elke cel uit een andere cel voortkwam ("Omnis cellula e cellula"). De functie van de celkern bleef lange tijd onduidelijk.[58]
Tussen 1877 en 1878 publiceerde Oscar Hertwig een reeks onderzoeken over de voortplanting van zee-egels, waaruit bleek dat de kern van de zaadcel tijdens de bevruchting versmelt met de kern van een eicel.[59] Voor het eerst werd gesuggereerd dat een individu zich ontwikkelt vanuit een (enkele) celkern, wat veel discussie teweegbracht. Hertwig bevestigde zijn bevindingen in andere diergroepen, waaronder amfibieën en weekdieren. In 1884 kwam ook Eduard Strasburger tot dezelfde conclusie voor de bevruchting in planten. Vanaf die periode werd erfelijkheid meer en meer met de celkern in verband gebracht.
De functie van de celkern als drager van genetische informatie werd pas duidelijk nadat de mitose werd ontdekt en deze nieuwe kennis aan het begin van de 20e eeuw samengevoegd werd met de wetten van Mendel (chromosoom-erfelijkheidstheorie).[58] Na de uitvinding van de elektronenmicroscoop en fluorescentiemicroscoop vanaf de jaren 30 konden structuur en dynamiek van de celkern in groot detail worden beschreven.
Zie ook
Noten
Bronnen
Dit artikel of een eerdere versie ervan is een (gedeeltelijke) vertaling van het artikel Cell nucleus op de Engelstalige Wikipedia, dat onder de licentie Creative Commons Naamsvermelding/Gelijk delen valt. Zie de bewerkingsgeschiedenis aldaar. Literatuur
Externe links
|
{kind=link}
| Celorganellen van eukaryoten | ||||||
|---|---|---|---|---|---|---|
|
| Zie de categorie Cell nucleus van Wikimedia Commons voor mediabestanden over dit onderwerp. |