Nucleosoom
Een nucleosoom is een complex van DNA en histonen dat de genexpressie regelt. Nucleosomen zijn de met een elektronenmicroscoop zichtbare "kralen" in een chromatineketen. Het zijn de plaatsen waar de DNA-streng zich om de histonen windt. De nucleosomen op hun beurt liggen als een spiraal gewonden en vormen met acht nucleosomen per winding een structuur met een diameter van 30 nm.
| Portaal Genetica |
Opbouw is onvolledig weergegeven.
De nucleosomenhypothese werd voor het eerst in 1974 voorgesteld door Don en Ada Olins[1] en Roger Kornberg[2][3] en was een belangrijke stap voor het begrijpen van de genexpressie bij eukaryoten.
Het nucleosoom in de kern
Nucleosomen dienen twee belangrijke doelen in de celkern. Als eerste zorgen ze door het vormen van de kleinste samenballing (spiralisering), dat het dubbelstrengs DNA (dsDNA), dat ca, 2 meter lang is, in de celkern past. Daarnaast zijn ze belangrijk voor de werking van de transcriptie, doordat ze ervoor zorgen dat het RNA-polymerase zich niet hecht aan promotors van genen die voor de translatie op dat moment niet nodig zijn. Door verschuiving van een nucleosoom als gevolg van bepaalde enzymen, kunnen nieuwe genen afgelezen worden. Nucleosomen zijn ook de belangrijkste dragers van epigenetisch erfelijke informatie.
Structuur van de kern van het nucleosoom

Het nucleosoom omvat, met enige variatie en uitzonderingen, ruwweg 200 baseparen (bp) van het eukaryotische chromatine. De kern van het nucleosoom (blauw gekleurd in de afbeelding), die een doorsnede heeft van 11 nm en 5 nm hoog is, omvat ongeveer 147 bp dsDNA. Deze zijn gewikkeld in 1,67 linksdraaiende windingen om een kern van histon-eiwit. De andere ca. 10 - 80 bp, van het nucleosoom bestaat uit koppelings-DNA, vrij DNA, dat de kernen van elkaar scheidt. Formeel bestaat het nucleosoom uit de kern en eenzijdig het koppelings-DNA. Vaak echter wordt alleen de kern aangeduid als nucleosoom.
De meeste van de belangrijke eigenschappen waren al bekend in 1997 met de publicatie van de structuur van het nucleosoom, vastgesteld bij een resolutie van 2,8 Å.[4]. De kristalstructuur van het nucleosoom is met behulp van röntgendiffractie bij een resolutie van 1,9 Å in 2002 vastgesteld.[5].
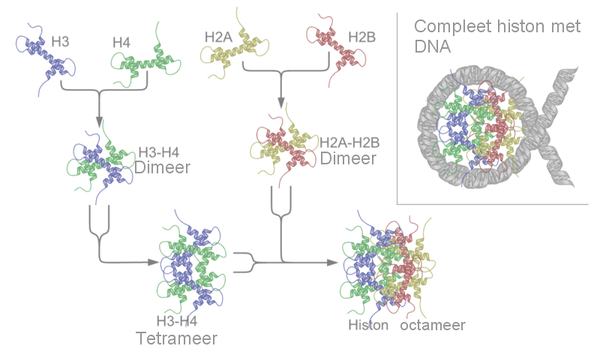
Elk van de vier histonen (H2A, H2B, H3 en H4) hebben een sterk op elkaar gelijkende structuur bestaande uit drie alfa-helices, die met lussen van elkaar gescheiden zijn. In een oplossing paren histonen met identieke kopieën en worden dimeren of histongevouwen paren genoemd. In het geval van de H3 en H4 histonen vormen de dimeren tetrameren door waterstofbruggen. Bij de vorming van een nucleosoomkern gaat eerst het H3-H4 tetrameer aan het dsDNA zitten, die daarna een verbinding aangaat met twee afzonderlijke H2A-H2B dimeren, die zich tegelijkertijd aan het tetrameer binden.
Het histon-octameer heeft ruwweg met elke 10 bp een wisselwerking met het om zich heen gewikkelde dsDNA. Elk van de vier histonen heeft drie wisselwerkingsregio's met het dsDNA. De centrale wisselwerkingsplaats van elk dimeer wordt door middel van een waterstofbrug gevormd door een α-helix van elk histon in het dimeer dat wijst naar een alleenstaande fosfaatgroep op het dsDNA. Aan beide zijden wordt op plaatsen, die 10 bp van elkaar zitten, door beide histonen een lus met behulp van een waterstofbrug met andere alleenstaande fosfaatgroepen gevormd. Twee andere wisselwerkingen van in totaal 14 vinden plaats door de wisselwerking van de staarten van elk van de H3 histonen. Deze vinden plaats op de ingangs- en uitgangspunten van het om het nucleosoom gewonden dsDNA, en helpen om deze plaatsen aan de kern van het nucleosoom vast te zetten.
Buiging van de superhelix gebeurt primair wanneer de kleine of de grote groef tegenover het octameer ligt en komt daarom voor in treden van ruwweg 5 bp. De buiging om het octameer bij de grote groef gaat gemakkelijk, terwijl de buiging bij de kleine groef gebeurt met behulp van de zijketens van arginine die in de groef steken. Bij het H3-H4-tetrameer gaat dit gemakkelijk, maar bij de H2A/H2B-dimeergebieden verloopt het met een knik.[6]
Vele eiwitten binden alleen aan specifieke DNA-sequenties (DNA-plaatsen). Hoewel nucleosomen een voorkeur hebben om zich te binden aan bepaalde DNA-sequenties, kunnen ze zich bijna aan elke DNA-sequentie binden. Het blijkt dat watermoleculen het aantal histon-DNA-wisselwerkingen ruwweg verdubbelt, doordat ze tussen de atomen gaan zitten die normaal te ver van elkaar liggen voor het vormen van waterstofbruggen.[5] Door deze flexibiliteit als gevolg van de watermoleculen kan een histon-octameer een zeer grote variatie aan DNA-sequenties om zich heen winden.
Structuur en doel van de histonstaarten
Het uiteinde van een histoneiwit heeft en staart van aminozuurresten met een lengte die afhankelijk is van het type histon. Het doel van de staart is op dit ogenblik nog niet geheel duidelijk, maar het schijnt dat ze bijdragen aan de stabiliteit van het nucleosoom[7] en als aankoppelplaats voor andere eiwitten. De structuur van de staart kan door enzymen in de celkern iets gewijzigd worden en dit speelt mogelijk een rol bij de vorming van hogere-orde-structuren van het chromatine.[8]
Nucleosoomvorming in vitro
Nucleosomen kunnen in vitro gemaakt worden door óf gebruik te maken van gezuiverde natuurlijke of recombinant-histonen óf met behulp van hun verschillende alternatieve structuren.[9]
De standaardtechniek maakt gebruik van zoutdialyse en berust op het feit dat een H3-H4-tetrameer bij hogere zoutconcentraties een verbinding aangaat met het DNA van het H2A/H2B-dimeer. Een reactie van vier kernhistonen met een naakt DNA-template (DNA-matrijs) wordt als eerste in gang gezet bij 4 °C en een zoutconcentratie van 2M. Deze zoutconcentratie wordt geleidelijk verlaagd door met de hand de buffer te vervangen of door een lagere zoutbuffer erbij te pompen en tegelijk dezelfde hoeveelheid eruit te pompen. Door het verlagen van de zoutconcentratie gaan ongebonden tetrameren in oplossing en verbinden zich daarna aan de DNA-matrijs. Bij nog verdere verlaging van de zoutconcentratie gaan de H2A/H2B-dimeren verbindingen aan met het tetrameer op de matrijs.
Referenties
|
| Het genoom | chromosoom · chromatine · chromatide · histonen · telomeer · centromeer · geslachtschromatine · nucleosoom |
| Chromosomen | karyogram · homologe chromosomen · isochromosoom · autosoom · heterosoom · geslachtschromosoom · X-chromosoom · Y-chromosoom |
| Ploïdie | diploïdisatie · haplogroep · haplotype · haploïdie · diploïdie · dihaploïdie · triploïdie · tetraploïdie · hexaploïdie · octoploïdie · polyploïdie · aneuploïdie · autoploïdie · alloploïdie · euploïdie |
| Genetica | crossing-over · recombinatie · fractionatie · syntenie · mutatie · genoommutatie · puntmutatie · segmentmutatie · basensubstitutie · frameshiftmutatie · deletie · duplicatie · indel · inversie · insertie · translocatie |
| Celdeling | mitose · meiose · karyogamie · spoelfiguur · centrosoom |