Gametofyt
Een gametofyt is, binnen de levenscyclus van planten en algen met geslachtelijke voortplanting, de generatie ('het stadium') die de nieuwe gameten vormt.
Als de gametofyt diploïde is, worden de gameten in gametangia gevormd door (gametische) meiose (reductiedeling), maar als de gametofyt haploïde is, worden de gameten door gewone celdeling, mitotisch gevormd. Dit laatste komt het meeste voor.
Levenscyclus

Veel organismen en met name planten (in wijdere zin) zijn diplohaplonten, dat wil zeggen dat er een afwisseling is van een haploïde gametofyt-fase met een diploïde sporofyt-fase[1][2]
Bij zaadplanten, varens en mossen is de gametofyt haploïde (1n): er is slechts enkele aantal chromosomen aanwezig. Bij de sporofyt is dit een dubbele chromosomenset: deze zijn diploïde. Bij de meiose worden eerst de homologe chromosomenparen van elkaar gescheiden, waarna ook beide chromatiden gescheiden worden. Deze planten zijn diplohaplonten: de generatie van de gametofyt wordt na de bevruchting afgewisseld door een diploïde generatie van de sporofyt.
Bij planten, algen en schimmels zijn de generaties vaak van een verschillende vorm (heteromorf), waarbij óf de gametofyt óf de sporofyt een forsere ontwikkeling en langere levensduur heeft. Men spreekt hier van een heteromorfe diplohaplont met dominante een gametofyt, respectievelijk met een dominante sporofyt. In dit laatste geval zijn de gametofyten nauwelijks meer zichtbaar en herkenbaar. Bij planten zijn er, ondanks de grote verscheidenheid in de generatiewisseling, toch duidelijke maar soms moeilijk herkenbare onderlinge overeenkomsten. Wilhelm Hofmeister onderkende halverwege de 19e eeuw als eerste de overeenkomst in de generatiewisseling binnen de Embryophyta van de mossen, varens en naaktzadigen.
Het groenwier zeesla is echter een voorbeeld van een generatiewisseling waarbij de sporofyt en gametofyt vrijwel gelijkvormig zijn: zeesla is een isomorfe diplohaplont.
Diplohaplonten bij Embryophyta
| Diplohaplont met digenetische cyclus | ||||||
| kernfasen | ||||||
| diplofase (2n) |
← → |
haplofase (1n) |
||||
| → Gene- raties → |
zygote | bevruch- ting ← |
gameten |
|||
| ↓ | ↑ | |||||
| gameto- fyt | ||||||
| (meio-) sporo- fyt |
↑ | |||||
| sporische → meiose |
spore | |||||
Embryophyta (ook wel "landplanten") zijn diplohaplonten, waarbij een kernfasewisseling optreedt met een haplofase (haploïde fase) en een diplofase (diploïde fase). De levenscyclus is digenetisch, wat wil zeggen dat er een generatiewisseling is met "om en om" een haploïde gametofyt en een diploïde sporofyt. De vorming van de sporen gaat gepaard met reductiedeling of meiose; dit heet sporische of intermediaire meiose. Hierbij ontstaan haploïde sporen, die ook wel meiosporen genoemd worden.
De kernfasewisseling bestaat afwisselend uit een haplofase, waarin de cellen haploïde zijn en een "enkele set" van chromosomen hebben (1n), en een diplofase, waarin de cellen diploïde zijn en een "dubbele set" van chromosomen hebben (2n).
In een sporangium van de diploïde sporofyt worden door meiose de haploïde sporen gevormd, die uitgroeien tot gametofyten. Deze sporen en de resulterende gametofyt zijn haploïde: ze hebben de "enkele set" van chromosomen. De gametofyten produceert mannelijke of vrouwelijke gameten door mitose: de zaadcellen en de eicellen. De samensmelting van een mannelijke en vrouwelijke gameet geeft een (diploïde, dus met een dubbele set chromosomen) zygote, die zich ontwikkelt tot een sporofyt.
Tot de Embryophyta behoren enerzijds de mossen en verwanten (levermossen, hauwmossen) die een dominante gametofyt hebben, en anderzijds de vaatplanten (varens, naakt- en bedektzadigen) die een dominante sporofyt hebben. De term "dominant" slaat op de sterkere van ontwikkeling en op langere levensduur. Bij de vaatplanten domineert de sporofyt en is de gametofyt, hoewel altijd aanwezig, soms nauwelijks waarneembaar. Vaak is het dan nodig anatomische preparaten te maken.
| Groepen ↓ |
geslachts- verdeling[3] |
Gametofyten bij Embryophyta: mossen, varens en zaadplanten | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| haploïde gametofyt-fase, haplofase | bevruch- ting[4] | |||||||||
| spore[5] | prothallium, voorkiem |
haploïde "plant" |
gametangia:[6] antheridia, archegonia |
gameet[7] | ||||||
| Marchantio- phyta levermossen |
isospoor[8] | ♂ | ♂ sporen, eerst in tetraden, met elateren[9] |
♂ protonema[10], vrijlevend, fotoautotroof[11] |
♂ mosplant, thalleus of folieus, fotoautotroof |
antheridia | biflagellate spermatozoïden |
interne oögamie[12] | ||
| ♀ | ♀ sporen, eerst in tetraden, met elateren |
♀ protonema, vrijlevend, fotoautotroof |
♀ mosplant, thalleus of folieus, fotoautotroof |
archegonia | eicel | |||||
| Bryophyta (blad)mossen |
isospoor[13], zelden zwak heterospoor |
⚥ | sporen, eerst in tetraden, soms trileet |
protonema: chloronema (fotoautotroof), caulonema |
mosplanten, folieus, fotoautotroof |
antheridia | biflagellate spermatozoïden |
interne oögamie | ||
| archegonia | eicel | |||||||||
| Anthoceroto- phyta hauwmossen |
isospoor | ⚥ | sporen, met pseudo- elateren |
protonema, vrijlevend, fotoautotroof |
mosplant, thalleus, fotoautotroof |
antheridia | biflagellate spermatozoïden |
interne oögamie | ||
| archegonia | eicel | |||||||||
| Lycopsida Lycopodium wolfsklauw |
isospoor | ⚥ | sporen | prothallium, vrijlevend, saprofytisch |
antheridia | biflagellate spermatozoïden |
interne oögamie | |||
| archegonia | eicel | |||||||||
| Lycopsida Selaginella Engels mos |
heterospoor, hetero- sporangiaat |
♂ | microsporen | microprothalliumcel, endospoor, fotoautotroof |
antheridia | biflagellate spermatozoïden |
interne oögamie | |||
| ♀ | 4 macrosporen (tetrade), vrijkomend |
macroprothallium, endospoor, coenocytisch[14], fotoautotroof |
archegonia | eicel | ||||||
| Equisetopsida Equisetum paarden- staarten |
vrijwel isospoor, functioneel heterospoor, isosporangiaat |
♂ | androsporen met elateren |
♂ prothallium, fotoautotroof |
antheridia | spermatozoïden met spiraal- band van cilia |
interne oögamie | |||
| ♀ (→⚥) |
gynosporen met elateren |
♀ prothallium, fotoautotroof |
archegonia (+ antheridia) |
eicel (+ spermatozoïden) | ||||||
| Polypodi- opsida lepto- sporangiate varens |
isospoor | ♂ | sporen (trileet of monoleet) |
prothallium, vrijlevend, foroautotroof |
antheridia kort gesteeld |
polyflagellate spermatozoïden |
interne oögamie | |||
| ♀ | archegonia iets verzonken |
eicel | ||||||||
| Salviniales Marsileaceae pilvaren- familie[15] |
heterospoor, hetero- sporangiaat |
♂ | microsporen | microprothallium, endospoor, fotoautotroof |
2 antheridia | polyflagellate spermatozoïden |
interne oögamie | |||
| ♀ | macrosporen, vrijkomend |
macroprothallium, endospoor, fotoautotroof |
1 archegonium | eicel | ||||||
| Cycadales palmvarens |
heterospoor, hetero- sporangiaat, tweehuizig |
♂ | microsporen (→ pollen)[16] |
microprothalliumcel (basale cel) antheridium cel (apicale cel) |
pollenbuiskern + generatieve cel |
steriele cel + spermatogene cel → 2 spermatozoïden spiraal korte flagellen |
bestuiving + interne oögamie | |||
| ♀ | 1 macrospore[17] (3† van tetrade), in nucellus[18] |
macroprothallium, coenocytisch, → cellulair primair endosperm |
2-veel archegonia, archegonium- kamer |
eicel | ||||||
| Ginkgoales, Japanse notenboom |
heterospoor, hetero- sporangiaat, tweehuizig |
♂ | microsporekern (→ pollen) |
1ste prothallium cel + 2de prothallium cel + antheridium celkern |
pollenbuiskern + generatieve cel |
steriele cel + spermatogene cel → 2 spermatozoïden spiraal korte flagellen |
bestuiving, na 4 mnd. interne oögamie | |||
| ♀ | 1 macrospore (3† van tetrade), in nucellus met pollenkamer |
macroprothallium, coenocytisch |
2(-3) archegonia, archegonium- kamer |
eicel | ||||||
| Coniferales coniferen |
heterospoor, hetero- sporangiaat, eenhuizig |
♂ | microsporen (→ pollen met 2 luchtblazen) |
(1ste prothallium cel + 2de prothallium cel + antheridium celkern) |
pollenbuiskern + generatieve cel |
steriele cel + spermatogene cel → 2 ♂ gameten |
bestuiving; na enkele wk. tot 12 mnd.: sifonogamie | |||
| ♀ | 1 macrospore (3† van tetrade), in nucellus |
macroprothallium, coenocytisch |
(1-)2(- ca. 200) archegonia |
eicel | ||||||
| Gnetales, Gnetum |
heterospoor, hetero- sporangiaat, tweehuizig |
♂ | microspore → pollen (3-kernig) |
pollenbuiskern + generatieve cel |
steriele cel + spermatogene cellen → 2 ♂ gameten |
bestuiving + sifonogamie · dubbele bevruchting ... | ||||
| ♀ | "embryozak" 4 kernig (tetrade) |
"embryozak", coenocytisch macroprothallium |
ø (geen ! archegonia) |
eikernen (+ endosperm) | ||||||
| Gnetales, Ephedra |
heterospoor, hetero- sporangiaat, tweehuizig |
♂ | microspore → pollen |
1ste + 2de prothalliumcel antheridiumcelkern |
pollenbuiskern + generatieve cel |
steriele cel + spermatogene cellen → 2 ♂ gameten |
bestuiving + sifonogamie dubbele bevruchting | |||
| ♀ | 1 macrospore (3† van tetrade), in nucellus |
vrije kerndelingen → cellulair prothallium |
2 - 3 archegonia | eicel | ||||||
| ventrale kanaalcel | ||||||||||
| Angio- spermae bedekt- zadigen |
heterospoor, hetero- sporangiaat, een- of tweehuizig |
♂ | microspore → pollen |
vegetatieve cel + generatieve kern → | 2 spermakernen | bestuiving + sifonogamie | ||||
| ♀ | macrospore binnen nucellus |
embryozak met antipoden |
synergiden | eicel/eikern | ||||||
| 2 polaire kernen |
secundaire embryozakkern (2n) |
dubbele bevruchting: secundair endosperm (3n) | ||||||||
| ||||||||||
Mossen, levermossen, hauwmossen
Mossen, levermossen en hauwmossen zijn organisme met een afwisseling van een sporofyt- en een gametofyt-generatie, waarbij de laatste domineert: de gametofyt is sterker ontwikkeld en heeft een langere levensduur dan de sporofyt.
| Heteromorfe diplohaplont met dominante gametofyt[1] | ||||||||||
| kernfase | ||||||||||
| diplofase (2n) |
← → |
haplofase (1n) |
||||||||
| Ge- ne- ra- ties |
zygote |
|
♂ gameten ♀ gameten |
|||||||
| ↓ | ↑↑ | |||||||||
| GAMETO- FYT | ||||||||||
| (meio-) sporo- fyt |
↑ | |||||||||
|
spore | |||||||||
De gametofytfase begint met een door meiose gevormde spore, die zich, via het stadium van de protonema (of voorkiem), ontwikkelt tot de gametofyt: het mosplantje.
Mossen zijn in het algemeen isospoor: de sporen zijn van gelijke grootte, uit de sporen groeien één- of tweehuizige gametofyten. Zo zijn bij parapluutjesmos (Marchantia polymorpha) de mannelijke en vrouwelijke gametofyten verschillend. Bij sommige mossen is er zelfs heterosporie: er zijn grote, vrouwelijke sporen die uitgroeien tot vrouwelijke mosplanten, en kleinere mannelijke sporen die zich ontwikkelen tot wat kleinere mannelijke mosplanten (deze worden wel "dwergmannetjes" genoemd) die groeien op de vrouwelijke planten.
_0559.jpg) Een bebladerd levermos (Lophocolea heterophylla).
Een bebladerd levermos (Lophocolea heterophylla). Het thalleuze levermos parapluutjesmos (Marchantia polymorpha) met paraplu-vormige sporangioforen.
Het thalleuze levermos parapluutjesmos (Marchantia polymorpha) met paraplu-vormige sporangioforen._9729.jpg) Een mos (Plagiomnium affine), stengeltje met blaadjes.
Een mos (Plagiomnium affine), stengeltje met blaadjes. Een hauwmos (Phaeoceros laevis) met antheridia en enkele jonge sporenkapsels op het thallus.
Een hauwmos (Phaeoceros laevis) met antheridia en enkele jonge sporenkapsels op het thallus.
De antheridia zij kortgesteelde bolvormige meercellige structuren met een uit steriele cellen bestaande wand en in het midden de zaadcellen.
De archegonia zijn flesvormige structuren met een lange hals, die nauwelijks gesteeld op de mosplant staan. De wand bestaat uit steriele cellen en in de buik bevindt zich een eicel. Na de bevruchting ontwikkelt zich de diploïde zygote zich tot een sporenkapsel (sporofyt) dat parasiteert op de gametofyt.
De gametofyt van levermossen zijn bebladerd (folieus) of thalleus. Bij thalleuze levermossen kan het thallus eenvoudig zijn van enkele cellagen dik toy gecompliceerd met luchtkamers en ademopeningen die op huidmondjes lijken. De folieuze levermossen zijn eenvoudiger gebouwd met stengeltje en hoogstens 1 cellaag dikke blaadjes zonder bladnerf. De blaadjes zijn meestal tweetoppig of tweelobbig en staan in drie rijen aan de opstijgende of liggende stengel: 1 rij kleinere buikblaadjes (amfigastria) en 2 rijen bovenblaadjes.
Bij bladmossen bestaat de gametofyt allereerst uit een meestal kortlevend, draadvormige protonema, waarop een groot aantal mosknoppen kan uitgroeien tot plantjes (die genetisch identiek zijn). Een mosplantje bestaat uit een stengeltje met de bladeren verspreid (in spiralen of in 5 rijen) langs de stengel en zijtakken. De bladeren hebben geen nerven, één nerf of twee korte. Nerven zijn meer cellen dik. Onder andere de veenmossen hebben geen bladnerf.
De eenvoudig gebouwde gametofyt van de hauwmossen lijkt op een thalleus levermos. De groene cellen van het thallus hebben een zeer afwijkende bladgroenkorrel die is voorzien van een pyrenoïde. Dit komt verder niet voor bij de landplanten, maar slecht bij enkele groepen van algen. Archegonia en antheridia zijn enigszins verzonken in het thallus.
Varens en varenachtigen
Wolfsklauwen
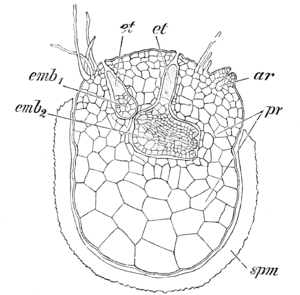
Van de in het Carboon belangrijke groep van de Lycopsida met door microfyllen gekenmerkte, ook vaak boomvormige soorten zijn er nog twee recente groepen van kruiden: Lycopodium en verwanten (wolfsklauw) en de Selaginella (Engels mos).
De eerste groep, de Lycopodiaceae, zijn isospoor, waaruit een fotoautotrofe of saprofytische of mycotrofe gametofyt groeit met ingezonken antheridia en archegonia.
De Selaginellaceae zijn heterospoor, dit in tegenstelling tot de Lycopodiaceae die isospoor zijn. Ook in de sporofytkenmerken verschilt Selaginella duidelijk van Lycopodium. De micro- en de macrosporen ontwikkelen zich in respectievelijk de micro en de macrosporangia, die in dezelfde strobilus staan. De sporofyten zijn vergaand gereduceerd: de sporen ontwikkelen zich al binnen de sporenwand, terwijl deze zich nog in het sporangium bevinden. Bij de bevruchting van de eicel door de antherozoïden kan de ontwikkeling van de jonge sporofyt soms al plaatsvinden binnen het sporangium.
Paardestaarten
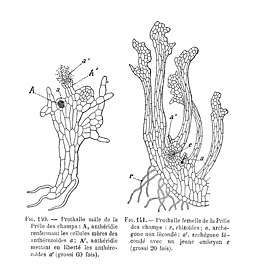
De Sphenopsida of Equisetopsida hadden evenals de Lycopsida in het Carboon een bloeiperiode met ook boomvormige soorten, terwijl en nu nog alleen kruidachtige vertegenwoordigers zijn.
Bij paardestaarten groeit de gekiemde spore uit tot een onregelmatig gevormd, betrekkelijk kleinblijvend, groen prothallium (of voorkiem). Het blijkt dat de helft van de sporen mannelijke prothallia oplevert, de andere helft groeit uit tot vrouwelijke prothallia. De antheridia zijn verzonken in het thallus en ook de archegonia liggen wat verdiept en hebben een kortere hals dan bij de mossen. Na de bevruchting groeit de nog binnen het archegonium liggende zygote uit tot een uit een spruit en een wortel bestaande sporofyt, die zelfstandig gaat leven.
Varens

Bij varens groeit de gekiemde haploïde spore uit tot een betrekkelijk kleinblijvend, groen prothallium. Het prothallium is vaak tot één cm groot, één tot enkele cellen dik, hartvormig of onregelmatig, kortlevend en lijkt uiterlijk op een thalleus levermos. De gametofyt vormt de geslachtsorganen. De archegonia en antheridia staan onder het prothallium enigszins verzonken in het thallus. Bij isospore varens is dit prothallium eenhuizig, groen en leeft vrij en zelfstandig zoals bij de eikvaren en de niervaren. Hierop groeien antheridia en archegonia.
Sommige varens zijn heterospoor: er is er een verschil tussen de mannelijke microsporen en de vrouwelijke mega- of macrosporen. De microspore ontwikkelt zich verder binnen de sporewand tot een microprothallium en één of enkele antheridia, die de zaadcellen vormen. Bij de macrospore vindt de ontwikkeling plaats binnen de sporewand tot een macro- of macroprothallium met één of enkele archegonia met elk een eicel. De "watervarens" zoals de vlotvarens, kroosvarens en Marsilea zijn een voorbeeld van varens met mannelijke en vrouwelijke prothallia.
Naaktzadigen
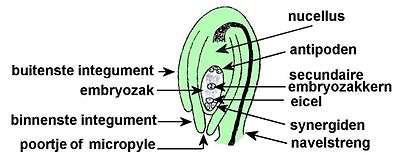
Ook bij zaadplanten (Spermatophyta, ook wel Fanerogamae, met de naakt- en bedektzadigen) worden gekenmerkt door een zaadknop of ovulum dat uitgroeit tot een zaad. Zaadplanten zijn heterospore vaatplanten met een endospore ontwikkeling van de macrogametofyt, waar de bevruchting plaatsvindt binnen de macrosporewand. De ontwikkeling van de gametofyt begint bij de micro- en de macrosporen. De sporen worden gevormd in de sporangia.
Een ovulum of zaadknop is een nucellus of macrosporangium, omgeven door één of twee omhullende integumenten (zaadvliezen). De macrosporen zijn vaak sterk gereduceerd: er worden in aanleg slechts 4 macrosporen gevormd in het macrosporangium. Van de 4 macrosporen gaan er meestal 3 te gronde, de vierde ontwikkelt zich verder tot de macrogametofyt. Onder "embryozak" of "kiemzak" kan zowel het macrosporangium als de daar gevormde macrospore worden verstaan - deze termen zijn dus vaag.[2] Bij naaktzadigen bevindt zich in de nucellus een pollenkamer, waar de stuifmeel- of pollenkorrels kunnen kiemen. De nucellus is omgeven door beschermende integumenten die aan het uiteinde de micropyle openlaten, waaruit bij sommige naaktzadigen een pollinatiedruppel kan uittreden, die pollenkorrels kan opvangen en bij uitdroging naar binnen zuigen. Bij bedektzadigen blijven de pollenkorrels op de stempel. Een of meer zaadknoppen staan op een vruchtblad of -schub.
De microsporen worden vertegenwoordigd door de pollenkorrels, die gevormd worden in de helmhokjes van de meeldraden (micrsporangia). De pollenkorrels komen vrij en worden door de wind, door dieren of op andere manier getransporteerd, zodat ze terechtkomen bij de vrouwelijk geslachtsorganen. De pollenkorrels van naaktzadigen hebben bij rijpheid niet meer het trilete of monolete litteken van hun ontstaan in tetraden, maar hebben een dunnere plaats in de sporewand, waar bij kieming de pollenbuis naar buiten kan groeien. Er ontwikkelt zich binnen de sporewand een zeer sterk gereduceerde microgametofyt, soms slechts bestaande uit een of enkele cellen of celkernen, zoals de antheridiumcellen, de pollenbuiskernen en een of twee generatieve kernen. Uiteindelijk zijn er twee spermatozoïden op spermakernen, waarvan er een de eicel bevrucht.
De pollenkorrel kiemt in zaadknop of op de stempel. Bij enkele naaktzadigen hecht het microprothallium zich met de pollenbuis vast en worden zaadcellen losgelaten in de pollenkamer, bij andere zaadplanten worden geen zaadcellen meer gevormd, maar brengt de pollenbuis de spermakernen naar de eicel.
Tot de naaktzadigen behoren onder andere de zaadvarens, de Cycadophyta, cycadofyten of palmvarens met twee of drie families (Cycadaceae en Zamiaceae, eventueel apart nog de Stangeriaceae), de Gnetophyta (ook wel als orde Gnetales, met de geslachten Gnetum, Ephedra en Welwitschia), de Ginkgophyta met slechts een recente soort: Ginkgo biloba of Japanse notenboom

Cycadofyten
Cycadophyta of "palmvarens", waartoe Cycas en Zamia behoren, zijn groenblijvende, tweehuizige planten met grote, geveerde bladeren. Ze zijn heterospoor, heterosporangiaat en tweehuizig. De sporofyllen staan in mannelijke en vrouwelijke kegels, maar bij het geslacht Cycas staan de macrosporofyllen aan de stam in kransen tussen de gewone macrofyllen.
De microsporen ontwikkelen zich in de microsporangia aan de microsporofyllen tot pollen. Men spreekt pas van pollen als de microsporen zich beginnen te ontwikkelen tot een meercellige microgametofyt. Deze vorm heeft een microprothalliumcel, pollenbuiscel, generatieve cel en antherozoïden met spiraalband van korte flagellen voor de bestuiving en de bevruchting.
De macrosporofyllen dragen twee of meer zaadknoppen met daarin een macrosporangium (nucellus). De daarin liggende macrosporemoedercellen ondergaan meiose en vormen daarbij elk 4 macrosporen die op een rij liggen in de nucellus. De verst van de micropile gelegen macrospore ontwikkelt zich verder tot macroprothallium, de overige 3 macrosporen gaan te gronde. Het macroprothallium is coenocytisch (het bestaat uit meer cellen met kernen maar zonder celwanden), en het vormt het primair endosperm met 2 tot veel archegonia, ieder met een archegoniumkamer.
Ginkgo
De tweehuizige Ginkgo heeft de microsporangia in paren op korte asjes ("sporangioforen" die op meeldraden lijken) ingeplant in strobili. De microsporangia ontwikkelen zich reeds voor de bestuiving tot pollenkorrels, waarin zich prothalliumcellen, een pollenbuiscel en een generatieve cel zich bevinden. Na de bestuiving (de pollenkorrel bevindt zich dan in de pollenkamer, een ruimte in de top van de nucellus) deelt zich de generatieve cel is een steelcel en een spermatogene cel. De pollenbuis groeit uit en dringt binnen in het nucellusweefsel. Later deelt de spermatogene cel zich tot twee spermatozoïden met een spiraal van korte flagellen. Deze spermatozoïden kunnen nu in de pollenkamer naar de eicel zwemmen, waar ze deze bevruchten.
Ginkgo (Japanse notenboom) is heterospoor, heterosporangiaat en tweehuizig. Het pollen groeit uit de microsporen. Hierin worden gevormd de microprothalliumcellen, een pollenbuiscel, een generatieve cel, die antherozoïden met spiraalband van korte flagellen vormt. De bestuiving wordt gevolgd door de bevruchting. Er is 1 functionele macrospore (3† van tetrade) in nucellus. De macrospore groeit uit tot een macroprothallium, dat eerst coenocytisch is, heeft later 2 archegonia met een archegoniumkamer en met een eicel.
Coniferen
De Coniferae (coniferen) (met onder andere de geslachten den, taxus, jeneverbes, Podocarpus en Araucaria) behoren tot de naaktzadigen. Zij hebben eenslachtige voortplantingsstelsels, meestal mannelijke en vrouwelijke strobili die op een (eenhuizige) plant staan. Een zaaddragende strobilus wordt 'kegel' genoemd, bijvoorbeeld 'dennenkegel'. De zaadknoppen staan gewoonlijk in paren op een zaadschub, die weer in de oksel staat van een dekschub (ook wel "bractee" genoemd). Onder andere omdat de zaadschub in de oksel staat van een dekschub wordt de zaadschub beschouwd als geëvolueerd uit fertiele assen en niet als afgeleid van een blad. De zaadknop bevat een nucellus met een macrosporemoedercel. Deze laatste ondergaat meiose en vormt 4 macrosporen, waarvan slechts één zich ontwikkelt via een coenocytisch stadium (met veel kernen zonder eigen celwanden om een gezamenlijke vacuole) door vorming van tussenwanden tot een meercellige macrogametofyt met enkele archegonia. Het archegonium heeft enkele halscellen en een eicel.
De mannelijke (of microsporangiate) strobilus is eenvoudig gebouwd uit 'microsporofyllen' die in een spiraal loodrecht op de as van de strobilus zijn ingeplant. Meestal staan er twee, maar soms tot 15 microsporangia (bij Araucariaceae) op het sporofyl. De microsporen ontwikkelen zich binnen het sporangium al tot pollenkorrels met geen tot enkele prothalliumcellen, pollenbuiscel en een generatieve cel. De pollenkorrels zijn voorzien van luchtblazen, waardoor ze goed door de wind verspreid kunnen worden. Na de bestuiving deelt de generatieve cel weer verder tot een steelcel en een spermatogene cel, welke laatste de mannelijke gameten vormt: cellen zonder flagellen of naakte kernen zonder eigen protoplast.
Bij de bevruchting groeit de pollenbuis vast in de nucellus en verder naar de eicel, waar de spermakern de eicel bevrucht.
Coniferae coniferen zijn heterospoor, heterosporangiaat en eenhuizig. De pollenkorrels groeien uit microsporen en hebben 2 luchtblazen. Er zijn microprothalliumcellen, pollenbuiscel en 2 generatieve kernen. De bestuiving + sifonogamie.
De macrosporemoedercel vormt 1 macrospore en 3† in een tetrade in de nucellus. Het macroprothallium is eerst coenocytisch en krijgt 2 archegonia met een eicel.
Gnetales
De orde Gnetales kent drie families: de Gnetaceae, de Ephedraceae en de Welwitschiaceae. Deze families nemen in hun levenscyclus min of meer een tussenpositie in tussen de bedektzadigen en de overige naaktzadigen.
Gnetum
De soorten van het geslacht Gnetum zijn een tweehuizige bomen of lianen.
In de ovula vormen zich in de nucellus enige macrosporemoedercellen waarvan de kernen zich alle door meiose delen zonder dat er celwanden worden gevormd: zo ontstaan er coenocytische macospore-tetraden (dit wordt ook wel genoemd: 4-sporige embryozakken).
Uit de ca. 10 macrosporen groeien hoogstens drie macroprothallia, die veelkernig zijn maar geen archegonia vormen. Enkele van de kernen fungeren als eikernen en kunnen bevrucht worden.
Microsporen groeien uit tot 3-kernige pollenkorrels met pollenbuis en 2 generatieve kernen (gameet- of spermakernen); men spreekt hier ook van sifonogamie. Elke van de spermakernen kan een eikern bevruchten, maar uiteindelijk ontwikkelt zich slechts één embryo in het zaad.
Ephedra
De soorten van het geslacht Ephedra zijn kruidachtige planten.
Pollenkorrels ontwikkelen zich uit microsporen met microprothalliumcellen, een pollenbuiscel, en 2 generatieve kernen die de spermakernen leveren voor de
De pollenzakjes (microsporangium) van de in een strobilus staande meeldraden, vormen uit de pollenmoedercellen (microsporemoedrecellen) de microsporen, die eerst in tetraden liggen. Binnen de wand van de microspore deelt zich de cel in enkele prothalliumcellen, een spermatogene cel, een pollenbuiscel en een steelcel. De spermatogene cel vormt twee spermakernen die zorgen voor de bestuiving en de sifonogamie: de bevruchting van de eicel en van de halskanaalcel: er is hier een dubbele bevruchting.
Het macrosporangium, dat is omsloten door een integument dat aan het uiteinde een micropilaire buis vormt, bevat een macrosporemoedercel of macrosporocyt. Deze macrosporocyt ondergaat meiose en vormt vier macrosporen op een rij. De drie het dichtst bij de micropile gelegen macrosporen gaan te gronde, de meest centraal in de nucellus gelegen macrospore ontwikkelt zich tot een coenocytisch macroprothallium. Tijdens de verdere groei ontstaat een meercellig macroprothallium met twee archegonia met kernen van een eicel en van een halskanaalcel. Deze worden beide bevrucht door de twee spermakernen uit de pollenbuis: dit is een dubbele bevruchting.
Bedektzadigen
In het jonge helmhokje ondergaat de microsporemoedercel twee delingen (meiose) en vormt microsporen (in een tetrade): 4 stuifmeelkorrels, die zich meestal van elkaar scheiden maar soms in een samenhangend groepje blijven. De haploïde kern van de microspore levert een mannelijke gametofyt (microgametofyt) met een kleinere generatieve kern en een grotere pollenbuiskern. De mannelijke gametofyt is de stuifmeelkorrel in het helmhokje van de meeldraad. De stuifmeelkorrel vormt na kieming op de stempel een generatieve kern. De generatieve kern deelt zich in 2 spermakernen die zorgen voor een dubbele bevruchting.
Het macrosporangium van de bedektzadigen is de nucellus met daarin macrosporemoedercel (macrosporocyte). De ondergaat meiose en vormt 4 lineair gerangschikte macrosporen. Hiervan gaan 3 macrosporen te gronde en 1 macropore ontwikkelt verder tot macrogametofyt met nog een mitotische deling zodat er in het totaal 8 kernen in twee groepjes van 4 zijn; de groei gaat ten koste van nucellus wat nu endosperm wordt genoemd. Uit elk groepje verplaatst zich 1 kern naar het midden en vormen daar de polaire kernen. De 3 overgebleven kernen aan de zijde van micropile vormen celwanden: een eicel en twee synergiden. De drie overgebleven kernen aan de tegenoverliggende zijde vormen celwanden en vormen zo de drie antipoden. De twee polaire kernen vormen 2–kernige cel: de embryozak.
Bestuiving (pollinatie) is de overdracht van pollen naar het stigma van de stamper van de bloem. De pollenbuis groeit uit met een pollenbuiskern en twee spermakernen. De pollenbuis groeit in een synergide, de twee spermakernen komen erin en de synergide desintegreert. De bevruchting is dubbel: een spermakern gaat naar eicelkern en er vindt karyogamie plaats: en levert een diploïde kern, de zygote. De zygote groeit verder tot een embryo. De andere spermakern gaat naar centrale polaire cel en versmelt met de twee polaire celkernen (1n tot 14n) en levert een triploïde cel (2n tot 15n), het primair endosperm, wat mitotische delingen ondergaat en het endosperm vormt.
De integumenten van de zaadknop vormen de zaadmantel en de wand van het vruchtbeginsel (=ovarium) wordt een deel van de vrucht.
De vrouwelijke gametofyt is de macro- of megagametofyt in de zaadknop en wordt ook wel de embryozak genoemd.[3]
Isomorfe diplohaplonten
Bij sommige algen zien de sporofyt en de gametofyt er gelijk uit. Een voorbeeld van een dergelijke isomorfe diplohaplont is het groenwier zeesla.
Trigenetische cyclus
Bij sommige roodwieren met een trigenetische cyclus zijn er twee diploïde sporofyt-generaties (de carposporofyt en de tetrasporofyt) die afwisselen met één haploïde gametofyt. De gameten hebben geen zweepstaartjes.
Haplonten
| Haplont met monogenetische cyclus[1] | |||||||||
| kernfase | |||||||||
| diplofase (2n) |
← → |
haplofase (1n) |
|||||||
| Ge- ne- ra- tie |
zygote |
|
← ♂ gameten ←♀ gameten |
||||||
| ↑↑ | |||||||||
| ↓ | gameto- fyt |
||||||||
| ↑ | |||||||||
| ↘ |
|
spore | |||||||
Bij haplonten is er alleen een haploïde generatie en treedt na de vorming van de zygote door bevruchting de meiose op; dit heet zygotische meiose. De enige generatie is dus de haploïde gametofyt.
Een haplontische levenscyclus vindt men bij de meeste schimmels, bij sommige groenwieren zoals Chlamydomonas. Voorbeelden zijn verder te vinden bij veel algen (Dinophyta, diverse Heterokontophyta en Chlorophyta), slijmzwammen (Acrasiomycota) en bij bepaalde schimmels (Zygomycota en diverse Chytridiomycota en Ascomycota).
Diplonten
| Diplont met monogenetische cyclus[1] | ||||||||
| kernfase | ||||||||
| diplofase (2n) |
← → |
haplofase (1n) |
||||||
| Ge- ne- ra- tie |
zygote |
|
♂ gameten gameten | |||||
| ↓ | ↑ ↑ | |||||||
| gameto- fyt |
|
↗↗ | ||||||
Diplonten zijn een diploïde organismes, waarvan alleen de gameten haploïde zijn. De meiose vindt direct plaats vóór de bevruchting en dus bij de vorming van de gameten. Men spreekt hier ook wel van gametische meiose.
In enkele gevallen worden diploïde mitosporen gevormd door mitose bij een mitosporofyt. Uit deze mitosporen groeit een diploïde gametofyt, die in door meiose de gameten vormt. Een voorbeeld is hier het groenwier Cladophora glomerata.[4]
| Diplont met digenetische cyclus[1] | ||||||||
| kernfase | ||||||||
| diplofase (2n) |
← → |
haplofase (1n) |
||||||
| Ge- ne- ra- ties |
zygote |
|
♂ gameten ♀ gameten | |||||
| ↓ | ↑↑ | |||||||
| mito- sporofyt |
↑↑ | |||||||
| ↓ | ||||||||
| mitospore | ||||||||
| ↓ | ||||||||
| gameto- fyt |
|
↗↗ | ||||||
Bij diplonten ontwikkelt zich uit de zygote (het product van de bevruchting) een diploïde gametenvormende generatie. De gameten worden gevormd door meiose: dit heet dan ook gametische meiose. Dit komt ook voor bij dieren, waar de enige haploïde cellen de gameten zijn. Een monogenetische cyclus komt voor bij Heterokontophyta zoals diatomeeën en bruinwieren, bij sommige groenwieren, bij waterschimmels en bij sommige ascomyceten. Een digenetische cyclus komt voor bij het groenwier Cladophora glomerata
Bronnen, noten en/of referenties
Literatuur
| |||
| Beschrijvende plantkunde: terminologie voor levermossen, mossen en hauwmossen | ||||||
|---|---|---|---|---|---|---|
|
| Beschrijvende plantkunde: terminologie voor varens en varenachtigen | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
|
| Levensvorm, groeivorm: | boom · chamaefyt · eenjarige plant · epifyt · fanerofyt · geofyt · groeivorm · hapaxant · helofyt · hemikryptofyt · houtige plant · hydrofyt · kruidachtig · levensduur · levensvorm · meerjarige plant · monocarpisch · overblijvend kruid · overblijvende plant · struik · teloomtheorie · therofyt · tweejarige plant · vaste plant · waterplant |
| Wortel: | bijwortel · centrale cilinder · diktegroei · endodermis · exodermis · luchtwortel · penwortel · pericambium · pericykel · rhizodermis · rizoïde ·secundaire diktegroei · centrale cilinder · topmeristeem · wortel · wortelhaar · wortelmutsje · zijwortel |
| Stengel: | bast · cambium · centrale cilinder · concaulescentie · diktegroei · knoop · lenticel · metatopie · stekel · stele · stengel · tak · topmeristeem · stam · uitloper · vertakking · wortelstok |
| Blad: | ader · blad · bladgroen · chloroplast · bladkussen · bladmoes · bladnerf · bladschede · bladschijf · bladstand · bladsteel · bladvoet · catafyl · cladoprofyllum · chlorenchym · fyllotaxis · hoofdnerf · kokertje · ligula · nerf · nervatuur · prefoliatie · ptyxis · steunblaadje · tongetje · tuitje · vernatie · zaadlob · zijnerf |
| Bloem, gameet, spore: | androecium · androfoor · androgynofoor · anthofoor · anthere · anthotaxis · bijkelk · bloemstengel · bloeiwijze · bloemgestel · bloem · bloembodem · bloembekleedsel · bloemdek · bloemdekblad · bloemkroon · bloemstengel · bractee · calyx · carpel · carpofoor · caulis · connectivum · corolla · discus · epicalyx · filament · funiculus · gametofyt · gynoecium · gynofoor · helmbindsel · helmdraad · helmhokje · helmhokje · hoogteblad · hypanthium · hypsofyl · inflorescentie · integument · kegel · kelk · kelkblad · knopligging · kroon · kroonblad · macrospore · meeldraad · meeldraaddrager · microspore · navelstreng · nucellus · omwindsel · ovarium · ovulum · periant · perigoon · petaal · placenta · pollenbuis · receptaculum · schijf · schutblad · sepaal · sporangium · spore · sporofyl · sporophyllum · sporofyt · stamper · stamperdrager · stempel · stengel · stigma · stijl · stylopodium · stylus · strobilus · tepaal · theca · vruchtbeginsel · vruchtblad · zaadbeginsel · zaadknop · zaadknopkern · zaadknopkern · zaadlijst |
| Vrucht, zaad, kieming: | carpel · cotyl · cryptocotylair · embryo · endosperm · epigeïsch · fanerocotylair · hypogeïsch · integument · kieming · kiemopening · kiemwit · mierenbroodje · perisperm · pluimpje · schijnvrucht · vaatmerk · vrucht · vruchtbeginsel · vruchtblad · zaad · zaadhuid · zaadlijst · zaadlob · zygote |
| Morfologie & anatomie: | apoplast · blad · bladgroenkorrel · bladstand · bloeiwijze · bloem · boomkruin · celwand · chloroplast · collenchym · cortex · cuticula · eicel · epidermis · felleem · fellogeen · felloderm · fenologie · floëem · fytografie · gameet · gametofyt · groeivorm · haar · houtvat · huidmondje · hypodermis · intercellulair · kelk · klierhaar · bloemkroon · kurk · kurkcambium · kurkschors · levensduur · levensvorm · merg · meristeem · middenlamel · palissadeparenchym · parenchym · periderm · plantaardige cel · plastide · schors · sclereïde · sclerenchym · spermatozoïde · sponsparenchym · sporofyt · stam · steencel · stengel · stippel · symplast · tak · thallus · topmeristeem · trachee · tracheïde · tylose · vaatbundel · vacuole · vrucht · wortel · xyleem · zaad · zaadcel · zeefvat · zygote |
| Bijzondere plantkunde: | algologie · bryologie · dendrologie · fycologie · lichenologie · mycologie · pteridologie |
| Paleobotanie: | archeobotanie · dendrochronologie · fossiele planten · gyttja · palynologie · pollenzone · varens · veen |
| Plantenmorfologie & -anatomie: | beschrijvende plantkunde · adventief · apoplast · blad · bladgroenkorrel · bladstand · bloeiwijze · bloem · bloemkroon · boomkruin · celwand · chloroplast · collenchym · cortex · cuticula · eicel · epidermis · felleem · fellogeen · felloderm · fenologie · floëem · fytografie · gameet · gametofyt · groeivorm · haar · houtvat · huidmondje · hypodermis · intercellulair · intercellulaire ruimte · kelk · kroonblad · kurk · kurkcambium · kurkschors · levensduur · levensvorm · merg · meristeem · middenlamel · palissadeparenchym · parenchym · periderm · plantaardige cel · plastide · schors · sclereïde · sclerenchym · spermatozoïde · sponsparenchym · sporofyt · stam · steencel · stengel · stippel · symplast · tak · thallus · topmeristeem · trachee · tracheïde · tylose · vaatbundel · vacuole · vrucht · wortel · xyleem · zaad · zaadcel · zeefvat · zygote |
| Plantenfysiologie: | ademhaling · bladzuigkracht · evapotranspiratie · fotoperiodiciteit · fotosynthese · fototropie · fytochemie · gaswisseling · geotropie · heliotropisme · nastie · plantenfysiologie · plantenhormoon · rubisco · stikstoffixatie · stratificatie · transpiratie · turgordruk · vernalisatie · winterhard · worteldruk |
| Plantengeografie: | adventief · areaal · beschermingsstatus · bioom · endemisme · exoot · flora · floradistrict · floristiek · hoogtezonering · invasieve soort · Plantengeografie · status · stinsenplant · uitsterven · verspreidingsgebied |
| Plantensystematiek: | taxonomie · botanische nomenclatuur · APG I-systeem · APG II-systeem · APG III-systeem · APG IV-systeem · algen · botanische naam · cladistiek · Cormophyta · cryptogamen · classificatie · embryophyta · endosymbiontentheorie · endosymbiose · evolutie · fanerogamen · fylogenie · generatiewisseling · groenwieren · hauwmossen · kernfasewisseling · korstmossen · kranswieren · landplanten · levenscyclus · levermossen · mossen · roodalgen · varens · zaadplanten · zeewier |
| Vegetatiekunde & plantenoecologie: | abundantie · associatie · bedekking · biodiversiteit · biotoop · boomlaag · bos · Braun-Blanquet (methode) · broekbos · climaxvegetatie · clusteranalyse · concurrentie · constante soort · differentiërende soort · ecologische gradiënt · ecologische groep · Ellenberggetal · gemeenschapsgradiënt · grasland · heide · kensoort · kruidlaag · kwelder · minimumareaal · moeras · moslaag · ordinatie · pioniersoort · plantengemeenschap · potentieel natuurlijke vegetatie · presentie · regenwoud · relevé · ruigte · savanne · schor · steppe · struiklaag · struweel · successie · syntaxon · syntaxonomie · Tansley (methode) · toendra · tropisch regenwoud · trouw · veen · vegetatie · vegetatielaag · vegetatieopname · vegetatiestructuur · vegetatietype · vergrassing · verlanding |