Pterosauriërs
De Pterosauria zijn een uitgestorven groep vliegende reptielen die behoren tot de Archosauromorpha. De grootste vliegende dieren die ooit op Aarde hebben geleefd behoorden tot de pterosauriërs.
| Pterosauria Status: Uitgestorven, als fossiel bekend Fossiel voorkomen: Trias — Laat-Krijt | ||||||
|---|---|---|---|---|---|---|
 | ||||||
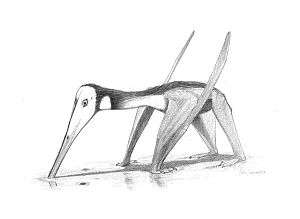
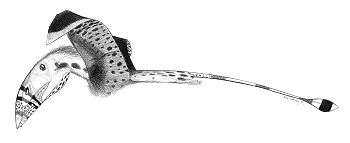
| "Coloborhynchus piscator" door John Conway geeft een typisch voorbeeld weer van een pterosauriër | ||||||
| Taxonomische indeling | ||||||
| ||||||
| Orde | ||||||
| Pterosauria Kaup, 1834 | ||||||
| Afbeeldingen Pterosauria op | ||||||
| Pterosauria op | ||||||
| ||||||
De pterosauriërs ontwikkelden zich meer dan 220 miljoen jaar geleden in het Trias en stierven 66 miljoen jaar geleden uit aan het eind van het Krijt. Ze waren de grootste dieren in de lucht, in de tijd dat de dinosauriërs de grootste dieren op het land waren. Zelf geen dinosauriërs, zijn ze er misschien wel nauw aan verwant. Hun afstamming is onzeker omdat er geen directe voorlopers van de pterosauriërs bekend zijn. Van de dieren die nu op Aarde leven, staan ze het dichtst bij de krokodillen en de vogels, maar die stammen niet van pterosauriërs af.
De pterosauriërs waren de eerste vliegende gewervelde dieren. Ze konden vliegen doordat de vierde vinger aan hun hand erg lang was en verbonden aan een soort vlieghuid zodat een vleugel gevormd werd. De vlieghuid bestond uit sterke vezels en spiertjes. Door die samen te trekken kon de vleugel op spanning gehouden worden en zijn welving, en daarmee het stijgvermogen, aangepast. De pterosauriërs waren hierdoor uitstekende vliegers. Hun borstkas was kort maar breed, voor een groot hart en goed ontwikkelde longen. Die longen waren verbonden met luchtzakken die zich vertakten tot in de botten, die op een dunne beenwand na hol waren. Het skelet was zo zeer licht maar toch sterk.
De pterosauriërs waren warmbloedig, waardoor ze heel actief konden en moesten zijn om veel voedsel te zoeken. Ze hadden schubben aan de voeten maar de rest van het lichaam was bedekt met een vacht van haren om warm te blijven. Voor de meeste bekende soorten bestond dat voedsel uit vissen en andere water- of zeedieren; sommige kleinere soorten leefden vermoedelijk van insecten.
De eerste pterosauriërs hadden een lange staart voor een stabiele vlucht. Op het eind van het Jura, 160 miljoen jaar geleden, ontwikkelde zich een deelgroep met een kortere staart, de Pterodactyloidea. Op de grond hadden die een meer opgerichte houding waardoor ze makkelijker konden lopen en opstijgen, en daardoor ook veel groter konden worden. Aan het eind van het Krijt hadden verschillende soorten een vleugelspanwijdte van over de tien meter. Dit waren de grootste vliegende dieren die ooit op Aarde hebben geleefd. Aan het eind van het Krijt werd de Aarde getroffen door een grote meteoriet en dat heeft vermoedelijk de laatste pterosauriërs uitgeroeid. In de ruim 150 miljoen jaar daarvoor moeten minstens enkele duizenden pterosauriërsoorten zijn geëvolueerd en weer uitgestorven.
Het eerste fossiel van een pterosauriër werd in 1784 ontdekt in Duitsland. Deze kreeg later de naam Pterodactylus ("vleugelvinger"). Pas langzaam begrepen de geleerden dat het om een uitgestorven reptiel ging in plaats van een vogel of een vleermuis. In 1834 werd de naam Pterosauria ("vleugelsauriërs") bedacht. In de negentiende eeuw werd nog een dozijn geslachten ontdekt en het feit dat het warmbloedige en behaarde dieren waren. Maar in het midden van de twintigste eeuw zag men ze als typische koudbloedige en primitieve reptielen. Vanaf 1970 echter hebben nieuwe vondsten het aantal bekende geslachten verzesvoudigd en tot het inzicht geleid dat ze toch warmbloedige en uitstekende vliegers waren.
Definitie
Een orde Pterosauria werd in 1834 benoemd door Johann Jakob Kaup.[1] Over de naam van die groep bleef eerst veel onzekerheid bestaan. Tot ver in de 20e eeuw was de aanduiding Ornithosauria gebruikelijker die in 1838 bedacht was door prins Karel Lucien Bonaparte.[2] Er was echter nooit enige twijfel over de inhoud: of een dier wel of niet tot de groep behoorde. De pterosauriërs delen namelijk een unieke bouw ofwel morfologie, die samenhangt met hun vermogen tot vliegen. Er zijn maar twee groepen Sauropsida bekend die echt kunnen vliegen. De ene, de vogels, bezit een vleugel die mede opgebouwd is uit veren. De andere groep zijn de pterosauriërs en hun vleugel heeft een geheel andere bouwwijze met een vlieghuid. Ondanks hun eigen bouw zijn de vogels daarmee niet strikt af te bakenen. Hun bekende soorten gaan zo geleidelijk over in andere vormen van de grotere groep waartoe ze behoren, de dinosauriërs, dat het volstrekt willekeurig wordt waar men de grens wil trekken. Bij de pterosauriërs daarentegen zijn geen fossiele overgangsvormen bekend die als een "missing link" kunnen dienen met niet-vliegende reptielen. Daarmee is hun groep tot nu toe ondubbelzinnig afgegrensd gebleven.
Het ontbreken van de schakels heeft voor de traditionele taxonomie, het opstellen van ontdekte levensvormen in een bepaalde ordening, dus voorlopig een heldere begripsvorming opgeleverd. De moderne evolutieleer echter wil juist alle levensvormen met elkaar in verband brengen door hun feitelijke stamboom vast te stellen. De methode van de cladistiek geeft daarbij van diergroepen een exacte wetenschappelijke definitie op basis van afstamming. Bij de keuze voor welke van de zeer vele op die wijze te bepalen groepen men de moeite wil nemen ze een naam te geven, speelt weer de bouw een rol. Het gebrek aan gegevens over de "ontbrekende schakels" zorgt bij het definiëren van de pterosauriërs dan voor grote problemen.
Een bepaalde clade (of klade), een groep die bestaat uit een voorouder en al zijn afstammelingen, kan men in de praktijk niet op de voor de hand liggende manier definiëren door meteen uit te gaan van de voorouder zelf:[3] die is meestal niet teruggevonden en hoe dan ook kan een directe afstammingsrelatie tussen soorten nooit echt bewezen worden. Eén alternatief is dat men een groep definieert als een verzameling van alle soorten die nauwer verwant zijn aan een bepaalde afstammeling dan aan een verwante soort die men juist van de groep wil uitsluiten. Zou men een dergelijke definitie gebruiken, bijvoorbeeld door de pterosauriërs te definiëren als alle soorten die nauwer verwant zijn aan een willekeurige bekende pterosauriër dan aan de vogels, dan is het vrijwel zeker dat in het begin van de afstammingslijn die naar de bekende pterosauriër voert, vormen blijken te zitten die qua bouw niet voldoen aan onze voorstelling van een lid van de Pterosauria en wel doordat ze helemaal niet kunnen vliegen. Dat gevaar kan men vermijden door in de definitie mede bepaalde gewenste nieuw geëvolueerde kenmerken (apomorfieën) op te nemen, in dit geval dus het vermogen tot vliegen.
Zo definieerde de paleontoloog Kevin Padian de klade Pterosauria als: alle Pterosauromorpha met een vierde middenhandsbeen en vinger die sterk uitgegroeid zijn om een vleugelmembraan te ondersteunen dat een synapomorfie is met Pterodactylus antiquus. Onder de pterosauromorfen, de groep van de pterosauriërs en hun directe verwanten, zouden dus alleen die soorten pterosauriërs zijn die het gedeelde nieuwe kenmerk (synapomorfie) van een grote ondersteunende vleugelvinger erfden van hun gezamenlijke voorouder, alsmede die voorouder zelf; Pterodactylus fungeert als willekeurige afstammeling. Dit soort definities is echter de laatste tijd impopulair geworden doordat ze nog steeds weinig exact zijn: wat is "sterk uitgegroeid" precies?[4]
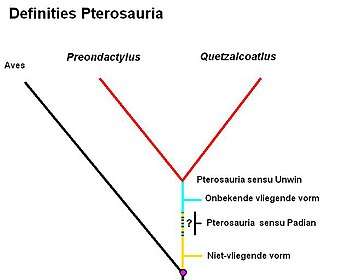
Om die reden is ook een tweede alternatief toegepast: de klade wordt gedefinieerd door uit te gaan van twee afstammelingen, de meest basale bekende soort (dat wil zeggen: de oudste bekende aftakking van de stamboom) en een willekeurige andere.[5] Zo definieerde amateurpaleontoloog David Peters de Pterosauria in 2000 als: de laatste gemeenschappelijke voorouder van Preondactylus buffarinii en Quetzalcoatlus northropi, en al zijn afstammelingen. Deze definitie werd overgenomen door de paleontoloog David Unwin. Hierbij werd ervan uitgegaan dat Preondactylus zich van alle bekende soorten het eerst heeft afgesplitst; als willekeurige andere afstammeling is de aansprekende soort Quetzalcoatlus genomen.[6] Paleontoloog Alexander Kellner gaf in 2003 een iets afwijkende definitie: de laatste gemeenschappelijke voorouder van de Anurognathidae, Preondactylus en Quetzalcoatlus, en al zijn afstammelingen, omdat zijn analyses aangaven dat de Anurognathidae wellicht nog basaler waren dan Preondactylus. Definities van het tweede type hebben het voordeel dat ze exacter zijn, en zeker niet te ruim: soorten zonder vliegende voorouder vallen erbuiten[7] — maar het nadeel dat ze juist te eng kunnen zijn. Ze kunnen nog onbekende basale vormen uitsluiten die in bouw overeenkomen met het klassieke beeld van een pterosauriër — namelijk als die zich eerder hebben afgesplitst dan de laatste gemeenschappelijke voorouder van de al bekende soorten. Mocht zo'n vliegend dier uit een eerdere aftakking alsnog opduiken, is de kans dus groot dat zulke definities worden aangepast. De prijs voor de exactheid is dus een zekere instabiliteit omdat de definitie niet weergeeft wat we eigenlijk bedoelen.
Evolutie
Afkomst
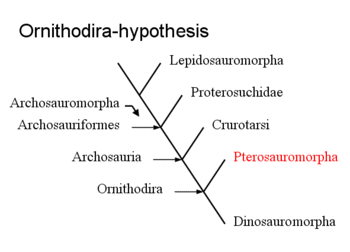
De oorsprong van de pterosauriërs is problematisch. Er zijn geen overgangsvormen bekend met andere groepen die ons ondubbelzinnig kunnen tonen waar hun directe verwantschappen liggen. Dit betekent ook dat zelfs de meest basale wel bekende pterosauriërs al zo sterk gespecialiseerd zijn, aangepast aan hun vliegende levenswijze, dat die evolutionaire verandering de sporen van hun afkomst voor een belangrijk deel heeft uitgewist. Wel is het met onze huidige kennis vrijwel zeker dat het diapside reptielen betreft — maar dat is een erg ruime afgrenzing want de meeste gewervelde landdieren zijn Diapsida. Een nadere bepaling kwam tot de jaren tachtig van de twintigste eeuw echter niet veel verder dan de hypothese dat ze vanwege het bezit van een schedelopening vóór de oogkas tot de archosauriërs zouden behoren, een groep die in de toenmalige betekenis min of meer overeenkwam met de huidige klade Archosauriformes. In dat decennium kwam de kladistiek op, een methode om verwantschap tussen groepen exact vast te stellen door te berekenen welke mogelijke stamboom het minste aantal evolutionaire veranderingen veronderstelt en dus het waarschijnlijkst is. Deze methodiek werd ook op het raadsel van de pterosauriëroorsprong toegepast en had als verrassende uitkomst dat de Pterosauromorpha, de pterosauriërs en hun directe verwanten, de zustergroep waren van de Dinosauromorpha, de dinosauriërs en hun directe verwanten, binnen een overkoepelende klade Ornithodira, zelf weer een deel van de Archosauria in moderne zin, een beperktere klade dan de Archosauriformes.


De meesten van de kleine groep traditionele wetenschappers die al op het gebied van de studie van pterosauriërs actief was, aanvaardden deze uitkomst niet maar ze werd steeds weer door nieuwe analyses bevestigd. In 1996 wees echter een van de auteurs van zo een analyse, Christopher Bennett, erop,[8] dat het bewijs voor deze "ornithodirahypothese" niet heel sterk was: de stamboom die ze oplevert was maar een paar stappen eenvoudiger (en dus maar iets waarschijnlijker) dan alternatieve stambomen die de Pterosauria basaal in (ten opzichte van de dinosauriërs dieper in de stamboom van) de Archosauromorpha (de ruimere groep die de Archosauriformes omvat) plaatsten. Daarbij kwam nog dat bijna alle veronderstelde synapomorfieën van de Ornithodira, dus de gedeelde nieuwe eigenschappen waarop het bewijs gebaseerd was dat de Pterosauromorpha en de Dinosauromorpha zustergroepen waren, zich in de achterpoten bevonden en te maken hadden met aanpassingen aan het enkelgewricht voor een versterkte afzet. Die eigenschappen zouden dus misschien helemaal niet duiden op een recente gemeenschappelijke afstamming en geen echte synapomorfieën zijn maar verklaard kunnen worden door convergente evolutie, het in niet nauw verwante groepen apart evolueren van schijnbaar gelijke kenmerken; wat betreft de pterosauriërs met name als een functionele aanpassing voor een verbeterd vermogen te springen, een zeer nuttige eigenschap voor vliegende dieren. Een volgend probleem was dat als pterosauriërs inderdaad de zustergroep van de Dinosauromorpha geweest waren, ze allerlei basale ("primitieve") en rudimentaire eigenschappen eerst verloren (want de laatste gemeenschappelijke voorouder van Archosauria had ze niet meer) en dan weer herwonnen (want zelf tonen ze die wel) moesten hebben, zonder dat daarvoor een goede functionele verklaring was. Ook deelden de Pterosauria geen enkele synapomorfie van de Archosauria; juist de nieuwe eigenschappen die op een gemeenschappelijke afstamming van de archosauriërs duiden, zouden bij de pterosauriërs toevallig allemaal weer verloren moeten zijn gegaan.
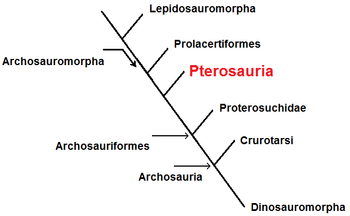
Bennetts conclusie was dat de pterosauriërs vermoedelijk geen nauwe verwanten van de dinosauriërs waren maar basale archosauroformen, iets dieper in de stamboom geplaatst dan de Erythrosuchidae of wellicht de Proterochampsidae. Meer verfijnde analyses zouden de beslissing moeten brengen in deze kwestie. De laatste jaren zijn zulke onderzoeken inderdaad uitgevoerd, maar ze spraken niet het verhoopte verlossende woord. De huidige situatie is in wezen gelijk aan die van de midden jaren negentig: de stamboom met de Pterosauromorpha als zustergroep van de Dinosauromorpha is de eenvoudigste maar slechts iets waarschijnlijker dan alternatieve oplossingen, zodat convergente evolutie een plausibele verklaring blijft. Invloedrijke onderzoekers als Peter Wellnhofer en David Unwin hebben de "ornithodira-hypothese" dan ook verworpen; Unwin nam aan dat de pterosauriërs basale archosauromorfen waren die zich vlak voor of (waarschijnlijker) na de Prolacertiformes hebben afgesplitst, die zelf zwevende vormen hebben voortgebracht als Sharovipteryx. Dat betekent niet dat ze nauwer aan de prolarcertiformen verwant zijn,[9] maar wel dat de archosauromorfe afstammingslijn op dit punt wellicht speciale kenmerken vertoonde die beide groepen gemeen hebben. Een enkeling, David Peters, veronderstelt zelfs dat het Lepidosauromorpha zijn, de grote groep waartoe onder andere ook de huidige hagedissen en slangen behoren, maar hij is daarin door niemand gevolgd.
Het gebrek aan overgangsvormen is deel van een algemener ontbreken van bekende Pterosauromorpha buiten de Pterosauria zelf. Er is maar één soort benoemd die — als de groep tenminste de zustergroep van de dinosauromorfen is[10] — een mogelijke basale pterosauromorf zou kunnen zijn en dat is Scleromochlus. De fossielen van deze vorm uit het late Trias (Carnien) zijn slecht bewaard gebleven en sommige analyses plaatsen hem als een zusterklade van de Ornithodira of een basale dinosauromorf, in plaats van een pterosauromorf. Verder zijn in 2002 nog "Pteromimus" en "Procoelosaurus" beschreven (maar niet officieel benoemd, vandaar de aanhalingstekens in plaats van het cursief) op grond van erg fragmentarische fossielen; deze nieuwe vondsten hebben tot nog toe weinig aandacht gekregen.

Scleromochlus is een klein dier met lange ledematen. Het kon ongetwijfeld redelijk klimmen maar had, voor zover we uit de schamele resten en vage afdrukken kunnen opmaken, geen speciale aanpassingen aan een levenswijze in de bomen, zoals sterk gekromde klauwtjes of gewrichten die armen en benen een groot grijpbereik gaven. Dit is direct relevant voor het vraagstuk langs welke evolutionaire weg de pterosauriërs hun vermogen tot vliegen verworven hebben. Net als bij het overeenkomstige probleem bij de vogels, zijn er twee hoofdscenario's: "omlaag" en "omhoog". Het model dat uitgaat van de beweging naar beneden, bijvoorbeeld vanaf een boom, heeft een groot verklarend vermogen: de bewegingsenergie van de val kan omgezet worden in een vlucht; de energie is verworven door omhoog te klimmen en dus is er een sterke selectiedruk die investering niet verloren te laten gaan door haar om te zetten in een voorwaartse beweging; het falen van die poging kan licht eindigen in de sterkste selectiefactor van allen: een onmiddellijke dood. Veel boombewonende diersoorten uit allerlei groepen hebben dan ook het vermogen tot glijden ontwikkeld. De overgang van een glijvlucht naar een slagvlucht is weliswaar biomechanisch zeer problematisch maar de vleermuizen, de derde groep echt vliegende gewervelden, lijken aan te tonen dat die tot stand gebracht kan worden. Anders dan vleermuizen maar net als dinosauriërs lijkt Scleromochlus met zijn lange poten echter meer een typische bodembewoner — althans naar de huidige inzichten: Friedrich von Huene die in 1914 als eerste een verband legde tussen Scleromochlus en de pterosauriërs, zag die poten nog als aanwijzing voor een leven in de bomen.[11] Veel onderzoekers menen dat de voorouders van de vogels door met hun armen te slaan, al hellingen oplopend of springend, het vermogen tot vliegen hebben verworven. Michael Benton stelde in 1999 de hypothese op dat een op Scleromochlus lijkende soort op dezelfde manier een vlieghuid zou hebben ontwikkeld om zijn sprongen op de grond te verlengen en zo de voorvader van de pterosauriërs werd.[12] Het probleem met een dergelijk "omhoog"-secenario is dat de selectiedruk in ieder geval veel zwakker is (veel voordeel kan de sprongverlenging niet geboden hebben en het is zelfs omstreden of het biomechanisch überhaupt zou werken) en de verklarende kracht en daarmee de kans dat het scenario het juiste is, dus kleiner. Moderne dieren die deze methode toepassen zijn niet bekend. Het traditionele alternatief als hypothetische voorouder, zoals voorgesteld door Rupert Wild in 1983, is dat van een boombewonend klein reptiel met ledematen in spreidstand, dat hij naar analogie met Gerhard Heilmanns vogelvoorloper "Proavis", "Propterosaurus" doopte en dat een vlieghuid ontwikkeld zou hebben om te glijden.[13] Een tussenoplossing is bedacht door Bennett: de voorouder zou door springen het vermogen tot vliegen ontwikkeld hebben — maar dan wel in een boom van tak tot tak.[14]
Een laatste vraagstuk in verband met hun afkomst is de ouderdom van de eerste pterosauriërs. De eerste bekende vormen duiken 217 miljoen jaar geleden op in het Norien. Als de "ornithodira-hypothese" juist is, kunnen de ornithodiren zich zo laat als het Ladinien, ongeveer 235 miljoen jaar geleden, afgesplitst hebben en leefde de eerste vliegende vorm misschien pas in het Carnien, circa 220 miljoen jaar geleden. Als pterosauriërs echter basale archosauromorfen zijn, moet hun afsplitsing veel eerder hebben plaatsgevonden en is het mogelijk dat de eerste pterosauriër al in het late Perm het luchtruim koos, meer dan 250 miljoen jaar geleden. In dat geval is er een lange verborgen afstammingslijn tijdens welke ze geen bekende resten hebben achtergelaten — vermeende vondsten van pterosauriërs uit het begin van het Trias bleken later altijd aan andere groepen toe te behoren — en dat betekent dat ze dan geen héél belangrijk deel van de fauna kunnen hebben uitgemaakt.
De eerste bekende pterosauriërs
De oudste bekende pterosauriër is Eudimorphodon, gevonden in het Norien van Italië; hij leefde 217 miljoen jaar geleden. Op dat moment moeten de pterosauriërs echter al een aanzienlijke diversificatie doorgemaakt hebben, want de meest basale vorm die we kennen — met mogelijke uitzondering van de in 2008 beschreven Raeticodactylus — is de iets later voorkomende Preondactylus en ook de Anurognathidae en de Dimorphodontidae die pas van vondsten uit het vroege Jura bekend zijn, moeten zich eerder hebben afgesplitst dan de groep waartoe Eudimorphodon behoort: de Campylognathoididae. Een mogelijke verklaring waarom er geen resten zijn die getuigen van hun eerdere evolutie, is dat de vroegste soorten bosbewoners waren: droge bossen leveren nauwelijks fossielen op. Pas toen sommige pterosauriërs boven zee op vis gingen jagen, zouden ze in de mariene sedimenten terecht zijn gekomen waarvan wij ze nu kennen. In deze periode waren de pterosauriërs zowel de enige vliegende gewervelden als, met een spanwijdte tot ongeveer twee meter, de grootste vliegende dieren; wat precies de interactie was met de insecten, de oudste en grootste groep vliegende dieren, en of ze ertoe bijgedragen hebben dat de allergrootste insecten verdwenen — er bestonden libellen met een spanwijdte van meer dan zestig centimeter — is nog onduidelijk.
De evolutie en verwantschap van deze vroege groepen is nog onzeker. David Unwin heeft een mogelijke ontwikkeling aangegeven op basis van exact kladistisch onderzoek: de meest basale vormen zoals Preondactylus, zouden nog relatief lange benen en korte armen gehad hebben en daarbij een oorspronkelijke, vrij hoge, schedelvorm. Een meer afgeleide bouw met krachtiger vleugels, zouden de overige pterosauriërs ontwikkeld hebben, samen de klade Macronychoptera vormend. Een aftakking daarvan, de Dimorphodontidae, behield de krachtige achterpoten maar de grootste tak, de Caelidracones, kreeg zwakkere benen. Die groep splitste zich weer in de Anurognathidae, die de oorspronkelijke bolle schedel behielden, en de Lonchognatha die een platte, lange schedel ontwikkelden. Die laatste groep splitste zich vervolgens in de Campylognathoididae en de Breviquartossa, die zich onderscheidden door meer subtiele schedel- en vleugelkenmerken. Al die groepen moeten dus al in het Norien aanwezig geweest zijn.
De Pterodactyloidea
Gedurende de loop van het Jura laten die eerst verborgen afstammingslijnen ons bekende fossiele resten achter; de pterosauriërs veranderen daarbij niet sterk van bouw. Het aantal ontdekte fossielen is overigens gering, onvoldoende om een goed beeld te krijgen van de ontwikkelingen. Uit het eind van het Jura echter zijn enkele rijke vindplaatsen bekend die als het ware een venster op hun evolutiegeschiedenis openen. Dan blijkt dat zich een groep heeft ontwikkeld met een sterk afwijkend Bauplan: de Pterodactyloidea. Terwijl eerdere pterosauriërs op vier poten zich vrijwel plat op de grond moeten hebben voortbewogen, met de vleugels ten dele gespreid, hadden de Pterodactyloidea op de armen steunend een sterk opgerichte houding, die mede mogelijk gemaakt werd doordat een veel groter deel van de vleugellengte gevormd werd door de middenhandsbeenderen, bij deze groep soms langer dan het opperarmbeen en onderarm samen. Aan deze houding was ook de stand van de kop aangepast die niet meer in het verlengde van een horizontaal gehouden nek lag maar haaks op de nu meer verticale nek stond. De staart, vroeger lang en een vluchtstabilisator vormend, is sterk gereduceerd en hetzelfde geldt voor de vijfde tenen die eerst een vlieghuid ondersteunden maar nu vrijwel verdwenen zijn. Wat de functionele verklaring kan zijn voor dit complex van veranderingen is nog erg onzeker. De traditionele interpretatie is dat ze hebben geleid tot een grotere wendbaarheid in de vlucht; de laatste tijd wordt de oorzaak eerder gezocht in een verbeterd loopvermogen om voedselbronnen op de grond te exploiteren, gecombineerd met een krachtiger afzet om sneller het luchtruim te kunnen kiezen en zo aan de op die grond voorkomende roofdieren te kunnen ontsnappen — de snelle en actieve Theropoda vormden een groot gevaar. Deze hypothese wordt ondersteund door het gegeven dat de twee deelgroepen van de Pterodactyloidea die uit het Jura teruggevonden zijn, de Dsungaripteroidea en de Ctenochasmatoidea, vormen hebben voortgebracht die door aanpassingen aan de tanden sterk gespecialiseerd waren in het voeden op de grond, door respectievelijk het kraken van schelpdieren en het filteren van kreeftjes uit het water.
Wat de afkomst van de Pterodactyloidea is, blijft nog onzeker. Volgens Unwin hebben ze zich binnen de Breviquartossa afgesplitst van de Rhamphorhynchidae, een in vorm wat meer ouderwetse groep die aan het eind van het Jura ook vele soorten telde. Hij sluit echter niet uit dat ze van de Anurognathidae een zustergroep zijn. In het verleden is weleens een poging gedaan die afkomst ook ruimtelijk te begrenzen door aan te nemen dat de Pterodactyloidea uit Europa zouden stammen en pas zich pas later naar andere continenten verbreid zouden hebben, maar ons gebrek aan kennis maakt het onmogelijk dit soort hypothesen serieus te onderbouwen. De mobiliteit van de pterosauriërs en de geringere afstand tussen de continenten in het Mesozoïcum staan ons niet toe hun in geologisch opzicht razendsnelle migraties te volgen. Ze kwamen ongetwijfeld op alle werelddelen voor en ook van een beperking tot lagere breedtegraden, zoals wel geopperd, bleek geen sprake.
In 1901 verdeelde Felix Plieninger, in een van de eerste pogingen hun interne relaties vast te stellen die nu nog gebruikte namen opleverde, de hele Pterosauria in twee onderorden: de Pterodactyloidea werden tegenover een "Rhamphorhynchoidea" gesteld, in welk laatste taxon alle "primitieve" vormen werden verenigd.[15] Dat de Pterodactyloidea niet de zustergroep van de Rhamphorhynchoidea vormen maar ervan afstammen, zou de Rhamphorhynchoidea echter parafyletisch maken: een groep die niet alle afstammelingen bevat. Dit soort begrippen is tegenwoordig in onbruik geraakt en dit geval laat al een reden zien waarom: de omvattende groep heeft dezelfde rang als de evolutionair gezien ingesloten groep.
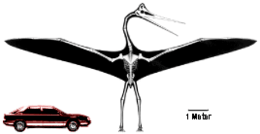
In het late Jura moeten zich nog twee andere deelgroepen binnen de Pterodactyloidea afgesplitst hebben: de Ornithocheiroidea, misschien wel de meest basale deelgroep, en de Azhdarchoidea. Beide zijn ons pas bekend door resten uit het vroege Krijt. Ze zouden, hoewel vermoedelijk niet nauw verwant,[16] allebei tandeloze vormen voortbrengen en aan het eind van die periode ook ware reuzen met een spanwijdte van over de tien meter; de verbeterde mobiliteit van de Pterodactyloidea, vooral wat betreft afzet en landing, maakte kennelijk een veel grotere omvang mogelijk.
Stamboom
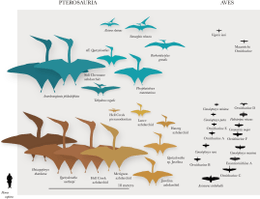
Over de onderlinge relaties van al deze groepen Pterosauria bestaat nog veel onzekerheid. Het is mogelijk het merendeel van de bekende en valide geslachten in een eenvoudige lijst onder te brengen die enigszins geordend is volgens een traditionele indeling langs suborden, superfamilies, families en onderfamilies (naar Unwin (2005)):
- I. "Rhamphorhynchoidea"
- Basale vormen (Preondactylus)
- Dimorphodontidae (Dimorphodon, Peteinosaurus)
- Anurognathidae (Anurognathus, Batrachognathus, Dendrorhynchoides, Jeholopterus)
- Campylognathoididae (Austriadactylus, Campylognathoides, Eudimorphodon)
- Rhamphorhynchidae
- II. Pterodactyloidea
- Ornithocheiroidea
- Ctenochasmatoidea
- Dsungaripteroidea
- Azhdarchoidea
- Lonchodectidae (Lonchodectes)
- Tapejaridae (Chaoyangopterus, Jidapterus, Sinopterus, Tapejara)
- Basale neoazhdarchiërs (Tupuxuara, Thalassodromeus)
- Azhdarchidae (Arambourgiania, Azhdarcho, Hatzegopteryx, Montanazhdarcho, Phosphatodraco, Quetzalcoatlus, Zhejiangopterus)
Zo een lijst geeft echter nog geen echte stamboom. Om niet ieder contact met de werkelijkheid te verliezen zijn er daarvan wel elementen ingevoegd: de "basale vormen" die niet bij een familie ingedeeld zijn en zelfs een verwijzing naar de klade Neoazhdarchia. Maar de verwantschap van de Pterodactyloidea is misleidend weergegeven omdat niet duidelijk wordt dat de directe zustergroep binnen de onnatuurlijke groep "Rhamphorhynchoidea" gezocht moet worden. Om een structuur te krijgen die de volle verwantschappen weergeeft en direct aansluit bij de resultaten van wetenschappelijk onderzoek, kunnen we een cladogram gebruiken zoals dit van Unwin:[17]
| Pterosauria |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Dit cladogram geeft echter slechts een van de vele cladistische analyses weer die de afgelopen jaren gedaan zijn. De eerste aangaande de interne verwantschappen van pterosauriërs werd in 1986 door Stafford C.B. Howse gepubliceerd.[18]
Bloei en uitsterven

In het vroege Krijt maakten de pterosauriërs hun bloeiperiode door; de meerderheid van de ons bekende vormen komt uit dit tijdperk en hun bouw toont dan de grootste diversiteit.[19] Het gaat dan om veel soorten Pterodactyloidea maar wellicht ook basalere soorten; de Anurognathidae en de Rhamphorhynchidae stierven misschien niet aan het eind van het Jura uit, zoals lange tijd gedacht werd. Wel is het zo dat de aantallen van die laatste groepen kennelijk afnamen. Misschien kwam dat door concurrentie met de Pterodactyloidea, maar het is ook mogelijk dat ze door een andere groep verdrongen werden, als voorbode van de neergang van alle Pterosauria: de vogels. De "oervogel" Archaeopteryx bewijst dat al in het late Jura de dinosauriërs vliegende vormen hadden voortgebracht. Als die een levenswijze hadden als boombewoner en insecteneter, moeten ze directe concurrenten geweest zijn voor de niches die veel pterosauriërs tot dan toe bezet hadden. De snelle diversificatie van de vogels in het Onder-Krijt vormt dus een plausibele verklaring voor althans een deel van de neergang van de basale vormen. In het Boven-Krijt worden die niet meer aangetroffen en helemaal op het eind van de periode, tegen het Maastrichtien, lijken er alleen nog maar grote tot reusachtige pterosauriërs over te zijn uit de Azhdarchidae en de Pteranodontia.
Dat er misschien nog maar twee deelgroepen bestonden, zegt op zichzelf niet zoveel — dat basale deelgroepen verdwijnen en vervangen worden door hun zustergroepen is een normaal proces, zonder noodzakelijke gevolgen voor de soortenrijkdom van de grotere groep als geheel — maar wel dat die maar een geringe variatie in levenswijze hadden. Vogels hadden toen ook viseters en watervogels voortgebracht en dit suggereert dat die de kleinere tegenhangers onder de Pterodactyloidea hebben doen uitsterven. Het wegvallen daarvan kan zeer wel het uitsterven van de hele Pterosauria veroorzaakt hebben want grotere soorten zijn veel sterker uitstervingsgevoelig doordat hun voortplanting trager verloopt (het kost meer tijd voordat individuen geslachtsrijp worden) en hun populaties kleiner zijn. Ze zijn daarom slechter in staat zich te herstellen van een terugval in aantallen. Dit zou bijzonder relevant hebben kunnen worden toen er zich 66 miljoen jaar geleden aan het eind van het Krijt, de Krijt-Paleogeengrens, een massa-extinctie voordeed, wellicht als gevolg van een meteorietinslag. Bij deze catastrofe die de meeste grote diersoorten doodde, zouden de kwetsbare reusachtige pterosauriërs zeker tot uitsterven gedoemd zijn.[20]
Het bovenstaande scenario is echter verre van zeker. Onze kennis van de pterosauriërevolutie is zo beperkt dat het heel goed mogelijk is dat het geleidelijk verdwijnen van de kleinere soorten een illusie is, veroorzaakt door toevallige variaties in conserveringsomstandigheden.[21] Ook is het niet duidelijk waarin de vogels eigenlijk een selectievoordeel hadden ten opzichte van de pterosauriërs. De hun vroeger wel toegeschreven superioriteit in vliegvermogen, stofwisseling en intelligentie bleek na beter onderzoek niet te hebben bestaan. Een poging dit probleem te kwantificeren had als uitkomst dat het effect toch reëel was aangezien anders ook slechte conserveringsomstandigheden meer fossielen hadden moeten opleveren, maar dat vogels de pterosauriërs niet zozeer verdrongen maar beter waren in het opvullen van de niches nadat een kleinere algemene uitstervingsgolf aan het einde van het Onder-Krijt het aantal soorten had verminderd.[22] Mochten de pterosauriërs echter in feite een grote vormenrijkdom genoten hebben tot aan het eind van het Krijt, dan hebben ze grote pech gehad want de gehele groep, groot of klein, lijkt hoe dan ook door de catastrofe te zijn uitgeroeid: pterosauriërs uit het Paleogeen zijn niet bekend. Van de soorten die tot dat moment geëvolueerd waren, is door ons ruim een honderdtal ontdekt. Als het scenario van een geleidelijke neergang door concurrentie met de vogels het juiste is, zal hun totaal minstens op enkele duizenden gelegen hebben.[23] Bleef de diversiteit tot het eind behouden, dan lag het nog enkele malen hoger. Er zijn echter geen duidelijke aanwijzingen voor een massale radiatie van erg kleine vormen, zoals die bij de zangvogels vele tienduizenden soorten heeft opgeleverd.[24]
Ontdekkingsgeschiedenis
Een schokkend fossiel: Pterodactylus
Er is wel beweerd dat oude legenden over draken geïnspireerd zouden kunnen zijn geweest door vondsten van pterosauriërfossielen. Maar hoewel pterosauriërs wel iets van een draak weghebben, zeker de tweevoetige verbeeldingen daarvan die populair waren tijdens de Middeleeuwen, zijn hun fossielen zo zeldzaam dat het hoogst onwaarschijnlijk is dat er enig oorzakelijk verband bestaat.
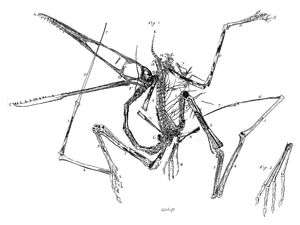
De wereld werd zich in 1784 met een schok van het bestaan van pterosauriërs bewust,[25] toen Cosimo Alessandro Collini, conservator van het naturaliënkabinet van keurvorst Karel Theodoor van Beieren, een artikel publiceerde over een uiterst wonderlijk fossiel waarop hij de hand had weten te leggen.[26] Het was een plaat fijnkorrelige kalksteen afkomstig van een groeve bij Eichstätt en toonde in uitstekende conservering het bijna volledige skelet van een op het oog volstrekt uniek dier.
Deze situatie stelde Collini voor een lastig probleem. Hij was een overgangsfiguur die vanuit zijn aanstelling nog fungeerde als een vroegmoderne geleerde die het puur te doen had moeten zijn om de (in dit geval zeer aanzienlijke) curiositeitswaarde — een eigenschap die nog in de zeventiende eeuw volledig serieus genomen werd. Aan de andere kant was hij, levende in de achttiende eeuw en ex-secretaris van Voltaire, een kind van de Verlichting, die zeer afwijzend stond tegenover alles wat "onverklaarbaar" leek. Het hele onderzoek naar fossielen, inclusief de fantasierijke interpretaties die aan hun bestaan gegeven werden, werd al gezien als een dubieus soort pseudowetenschap en iedere suggestie dat er monsters bestonden, was dus helemaal uit den boze — maar dit fossiel leek precies zo'n monster. Collini, die zag dat het wezen kennelijk vleugels had — bij veronderstelde dat de verlengde vingers een membraan ondersteunden — deed dus zijn best de vondst te duiden als een vogel of vleermuis, de twee "normale" groepen vliegende gewervelden, maar moest concluderen dat zo'n oplossing gewrongen was. De enige verklaring die hem overbleef, was dat het tegen de schijn in toch geen vliegend dier betrof maar een nog onbekend type zeedier.
Voor Collini was het daarbij niet vanzelfsprekend een "uitgestorven dier", want dat begrip zelf was in die tijd nog niet gangbaar. Soorten werden gezien als eeuwig en onveranderlijk en dat er een zou verdwijnen leek in strijd met de kosmische wereldorde of men die nu religieus of meer naturalistisch wilde interpreteren. Juist voor de toenmalige rationalistische natuurwetenschapper vergde het een enorme denkstap te aanvaarden dat onze huidige alledaagse werkelijkheid voorafgegaan was door een eerdere wereld van ontzaglijke ouderdom en bevolkt door "voorwereldlijke" wezens die nog het meest weghadden van fabeldieren. Onder invloed van enkele zeer overtuigende fossielen, waarvan dat van de pterosauriër het meest aansprekend was, vond in de generatie daarna toch een dergelijke omslag in het denken, ofwel paradigmaverschuiving, plaats. Belangrijkste vertegenwoordiger van die verandering was de Duits/Franse vergelijkend anatoom Georges Cuvier die het catastrofisme voorstond, de radicale leer dat de aarde tijdens haar lange geschiedenis met tussenpozen geheel of per continent verwoest was om dan weer door een scheppende macht bevolkt te worden. Over het meest exemplarische bewijs voor die leer, moest hij natuurlijk ook een oordeel geven. Cuvier zou later de reputatie krijgen op basis van een enkel botje een dier te kunnen determineren en met de vrijwel complete vondst van Collini had hij helemaal geen moeite: al in 1801 toonde hij in een korte samenvatting aan dat het evident een vliegend reptiel betrof; het cilindervormige os quadratum was daarvoor al genoeg bewijs.[27] In 1809 behandelde hij het fossiel in meer detail;[28] hij verwierp hierbij de theorie van Johann Friedrich Blumenbach dat het toch gewoon een watervogel was en benoemde een geslacht voor het dier: Ptéro-Dactyle ofwel "Vleugel-Vinger".

In 1809 had Cuvier het oorspronkelijke fossiel niet kunnen lokaliseren en had zich moeten baseren op, onder andere, een zeer exacte kopergravure die Collini in 1784 had laten vervaardigen door de beroemde graveur Egid Verhelst II, die daarmee 's-werelds eerste pterosauriërillustrator werd. Cuvier vreesde al dat het door het geweld van de napoleontische oorlogen verloren gegaan was maar in feite was het in München achtergehouden door Samuel Thomas von Soemmerring die er zelf een studie aan wilde wijden. Het resultaat daarvan bracht hij op 27 december 1810 naar buiten in een lezing, die hij in 1812 publiceerde.[29] Von Soemmerring gaf een eigen naam aan het dier: Ornithocephalus antiquus, de eerste binominale naam van een pterosauriër. Hij erkende dat het een voorwereldlijk dier betrof maar wilde het geen reptiel noemen. Het was volgens hem zowel een zoogdier, een vleermuis, als een tussenvorm tussen de zoogdieren en de vogels. Deze naar huidige begrippen tegenstrijdige constatering wordt begrijpelijker als we bedenken dat Von Soemmerring een "tussenvorm" niet opvatte als een dier dat in afstamming tussen twee groepen lag: evolutie was immers nog geen gangbaar begrip. Een tussenvorm was letterlijk iets wat een "vorm" vertegenwoordigde die tussen twee andere vormen in zat. Hij zei dus in wezen dat zijn Ornithocephalus qua vorm bij de zoogdieren hoorde maar van deze nog het meeste die van een vogel benaderde. Anatomisch gezien was Von Soemmerings studie weinig gelukkig: hij liet zich verwarren door de lange middenhandsbeenderen van de Pterodactyloidea en dacht dat het de onderarmen waren; de onderarmen zag hij weer aan voor de bovenarmen. Ook in de naamgeving zou hij goeddeels falen: hoewel de soortaanduiding gehandhaafd zou blijven, kreeg het geslacht door een aanpassing door Cuvier, in 1819 gepubliceerd door Lorenz Oken, de gangbare naam Pterodactylus.[30] Daarnaast diende Cuvier hem inhoudelijk van repliek: nog in 1812 toonde hij omstandig de fouten van Von Soemmering aan.[31] Daarbij gaf hij ook aan hoe hij de levenswijze van de vliegende reptielen zag. Ze zouden op hun achterpoten gelopen hebben, geleefd hebben van insecten en hun drie handklauwen gebruikt hebben om 's nachts in de bomen te hangen.
De Pterosauria krijgen hun naam

Von Soemmerring slaagde er echter wel in de meeste Duitse geleerden ervan te overtuigen dat het dier een zoogdier was. Toen Kaup in 1834 een orde Pterosauria benoemde,[32] als deel van de Amphibia (toen gezien als zowel de amfibieën als reptielen omvattend), was hij in zijn land een relatieve uitzondering. Dat "sauria", tegenwoordig gebruikt voor de hagedissen alleen en inderdaad afgeleid van het Klassiek Griekse saura of sauros, "hagedis", werd toen namelijk voor bijna alle uitgestorven reptielgroepen gebruikt en hij maakte zo een duidelijke keuze tegen Von Soemmerring door ze "Vleugelsauriërs" te noemen. Dat woord vond eerst dan ook geen algemene ingang hoewel het later wel overgenomen zou worden door de invloedrijke Richard Owen; zelfs degenen die het eens waren met de identificatie als reptielen gebruikten meestal de term Ornithosauria ("Vogelsauriërs") die prins Bonaparte in 1838 had bedacht.[33] Grote verwarring bleef daarbij bestaan over de precieze aard van de groep: in 1830 bracht Johann Georg Wagler ze samen met de ichthyosauriërs en de plesiosauriërs onder in de klasse der Gryphi als zeedier; hij meende dat de vingers enorme vinnen ondersteunden.[34] En nog in 1843 dacht de Brit Edward Newman dat het vliegende buideldieren waren.[35]
Gezien al die onzekerheid noemde men ze meestal maar gewoon "pterodactylen" en pterodactyles is in de engelstalige landen tot op de dag van vandaag als informele term gebruikelijker gebleven dan pterosaurs (of het correcter afgeleide pterosaurians). Die gewoonte lag des te meer voor de hand omdat Pterodactylus lange tijd het enige benoemde geslacht bleef waaraan alle verdere vondsten werden toegewezen. In 1817 had Von Soemmering een tweede fossiel beschreven als P. brevirostris, waarvan we nu weten dat het een jonge Pterodactylus antiquus betreft. In 1829 werd de eerste pterosauriër uit Engeland en het vroege Jura, in december 1828 gevonden door de beroemde fossielenverzamelaarster Mary Anning, benoemd door William Buckland: Dimorphodon, als P. macronyx.[36] In 1830 benoemde Carl Theodori Dorygnathus als P. banthensis. In 1831 beschreef Georg Augustus Goldfuss Scaphognathus (als P. crassirostris); hij meende op het fossiel zelfs sporen van pluimen of haren gezien te hebben.[37] In 1846 benoemde John Scott Bowerbank de toen grootste bekende pterosauriër Lonchodectes als P. giganteus. Nog in 1856 zag Friedrich August Quenstedt wat we sinds 1928 kennen als Campylognathoides als een P. liassicus, nadat hij een jaar eerder Cycnorhamphus als P. suevicus beschreven had. Dat was toen echter al wat ouderwets geworden want in 1846 had de "Vader van de Duitse paleontologie" Christian Erich Hermann von Meyer een tweede genus benoemd: Rhamphorhynchus, een hernoeming van Georg Graf zu Münsters P. longicaudus uit 1839. En eigenlijk had Von Meyer, zonder het te weten, al in 1833 een tweede geslacht een naam gegeven: Gnathosaurus, maar daarvan zou pas in 1951 duidelijk worden dat het een pterosauriër betrof. Zijn vierde genus zou in 1852 komen: Ctenochasma. In een grote monografie uit 1859-1860, Die Reptilien aus dem lithographischen Schiefer des Jura in Deutschland und Frankreich (welk boek met prachtige lithografieën was geïllustreerd, die met dezelfde kalksteen gedrukt waren die ook de beste fossielen had opgeleverd) waagde hij zich ook aan een eerste indeling van de pterosauriërs die echter al snel door de ontwikkelingen in de wetenschap achterhaald zou worden; zijn onderscheid tussen vormen met lange en korte staarten bleef echter nuttig. Von Meyer liet ook in zijn land het inzicht doordringen dat pterosauriërs reptielen waren.
Nieuwe inzichten en vondsten
Vanaf 1859 veroorzaakte Charles Darwins Origin of Species een revolutie in de biologie. De relaties tussen verschillende diergroepen werden daarna meestal geduid als een echte evolutionaire verwantschap, niet meer als een enkele overeenkomst in vorm alleen. Niet iedereen werd echter Darwinist. In Engeland wilde Richard Owen nog wel toegeven dat er wat variaties mogelijk waren op een bepaald bouwschema of archetype maar dat dieren radicaal van morfologie zouden kunnen veranderen, ontkende hij: pterosauriërs bijvoorbeeld zouden altijd koudbloedige reptielen geweest zijn en ook gebleven. Dat was nu juist wat de meest enthousiaste evolutionisten wilden ontkennen. St. George Jackson Mivart, eerst een van de meest felle voorvechters van de nieuwe leer, zag ze als een prachtige illustratie hoe de evolutie alle kaders kon doorbreken: hij meende dat ze de voorouders van de vogels waren.[38] Mivart, zelf geen expert op dit gebied, werd daarin bijgevallen door Harry Govier Seeley, die zich zou specialiseren in dit onderwerp. In 1864 benoemde hij ze als een onderklasse Saurornia om de relatie met vogels uit te drukken en stelde dat ze warmbloedig waren.[39] In 1870 publiceerde hij het eerste boek dat helemaal aan pterosauriërs gewijd was: Ornithosauria,[40] waarvan de titel al aangeeft dat hij nu Bonapartes term overgenomen had. Hoewel Seeley later met veel tegenzin zou toegeven dat de vogels toch niet van de pterosauriërs afstammen, bleef hij de laatsten zijn leven lang als hun nauwe verwanten zien, en die van de dinosauriërs en krokodillen. In 1901 publiceerde hij het eerste populair-wetenschappelijke boek ter wereld over pterosauriërs: Dragons of the Air.[41] Toen dat in 1967 herdrukt werd, was het nog steeds het enige in zijn soort. Seeley schetst hierin een beeld van de pterosauriërs dat nogal afwijkt van het typische moderne reptiel: niet alleen waren ze warmbloedig maar dus, ter isolatie, ook met haren bedekt en ze konden op twee poten rennend opstijgen.

Naast die theorievorming werden er ook weer nieuwe soorten gevonden. Seeley zelf benoemde in 1869 al Ornithocheirus, waarvan we nu weten dat het wellicht een van de grootste soorten omvat met een spanwijdte van tegen de tien meter. In die tijd was dat nog onbekend en daarom maakte in 1870 de eerste vondst uit Amerika grote indruk waar Othniel Charles Marsh in het kalkgesteente van Kansas grote skeletten opgroef die hij in 1872 beschreef als Pterodactylus ingens en P. occidentalis en in 1876 hernoemde tot Pteranodon; de fossielen wezen op een spanwijdte van acht meter. In hetzelfde jaar beschreef hij Nyctosaurus. Ondertussen had Owen in 1874 Coloborhynchus benoemd.
In 1882 werd er wat meer bekend over het uiterlijk van de pterosauriërs omdat toen zowel Marsh[42] als Karl Alfred Zittel[43] ieder een artikel publiceerden over een fossiel dat een afdruk toonde van het vleugelmembraan van Rhamphorhynchus. Opmerkelijk genoeg leidde Marsh uit zijn fossiel (in 1873 in Duitsland opgedolven en door hem voor $1000 aangekocht) af dat het membraan aan de enkels verbonden was, terwijl het toch net als Zittels vondst een zeer smalle vleugel toonde. Beiden verschilden van mening over de vraag of de vlieghuid ook aan de staart vastzat, zoals Marsh in navolging van Owen had aangenomen. Seeley koos in de controverse voor een model met vleugels die aan het middel vastzaten. Marshes fossiel had ook een vaantje aan het achtereind van de staart. Marsh reconstrueerde dat asymmetrisch als een richtingroer maar later werd het weer als een horizontaal hoogteroer gezien. In 1888 werd door Edward Newton het natuurlijk afgietsel van het brein van Parapsicephalus beschreven, dat toonde dat pterosauriërs vrij grote hersenen hadden.[44]
Het onderzoek valt stil
Al tegen het eind van de negentiende eeuw begon het onderzoek naar pterosauriërs te stagneren. Na 1876 werden er lange tijd geen namen gebaseerd op nieuwe vondsten toegevoegd aan het op dat moment bereikte totaal van veertien geslachten die nu nog als valide beschouwd worden, hoewel sommige al eerder bekende fossielen een nieuwe naam kregen. Nyctosaurus was de laatste van zijn eeuw en tot 1964 werden er maar twee echt nieuwe ontdekkingen beschreven: Anurognathus in 1923 en Batrachognathus in 1948. Sommige van de redenen voor deze stilstand waren heel specifiek zoals een gebrek aan goede en nog niet onderzochte vindplaatsen. Ook speelde een rol dat Seeley eigenlijk de enige specialist was op dit gebied; voor de andere wetenschappers vormde het maar een onbelangrijk deel van hun werkzaamheden. Hierdoor kwam geen institutionalisering tot stand; er was nergens een "leerstoel pterosauriërs" die de continuïteit van het onderzoek had kunnen garanderen.
Maar er was ook een meer algemene oorzaak: in de loop van de twintigste eeuw veranderde de hele paleontologie van karakter. Het nieuwe van de ontdekkingen was er af en men raakte, in plaats van de zoveelste soort te willen vinden, meer gericht op het leggen van grote verbanden zoals het assisteren van de geologie door het dateren en ordenen van aardlagen. De pterosauriërs waren door hun zeldzaamheid echter niet geschikt als gidsfossiel. Ook bleek de aanvankelijke werking van de evolutietheorie, dat door het aannemen van een afstammingsrelatie de bestaande denkbeelden over strikt afgegrensde diergroepen werden doorbroken, niet lang te duren: al snel keerde men terug naar de oude absolute concepten maar nu vermomd als een ordening binnen een evolutionaire vooruitgang. Het hele dierenrijk werd weer gerangschikt van laag naar hoog; op die ladder werden de pterosauriërs als "reptielen" geplaatst en Seeleys theorie dat ze warmbloedig en behaard waren, werd naar het rijk der fabelen verwezen. Als reptiel waren ze immers koudbloedig en geschubd. Door hun lage stofwisseling zouden ze niet echt hebben kunnen vliegen en de bouw van hun vleugels zou daarom primitief en inefficiënt geweest zijn. In het meest gebruikte paleontologische standaardwerk van het midden van de twintigste eeuw, Vertebrate Paleontology van Alfred Roemer, werd beweerd dat pterosauriërs hun vlieghuiden niet op spanning hadden kunnen houden; ze misten daarin immers de extra ondersteunende vingers die vleermuizen bezitten. Tijdens de neerwaartse slag zou het membraan zijn gaan wapperen waardoor er geen opwaartse kracht opgewekt zou kunnen worden en bij de vleugelopslag zou het vleugelprofiel zelfs helemaal omgeslagen zijn. De pterosauriërs waren zo gedoemd tot een neerwaartse glijvlucht die hoogstens door thermiek wat verlengd werd. De voortbeweging op het land zou al even onbeholpen geweest zijn. Met moeite zouden de beesten langzaam weer in een boom of op een klif geklauterd zijn om weer even door de lucht te kunnen glijden. Slechts een enkele keer per dag hadden ze ondertussen een insect of vis weten buit te maken — maar als koudbloedige dieren was hun energiebehoefte toch al niet groot. De pterosauriërs leefden ook in het "Tijdperk der Reptielen" en dat werd gezien als een tussenfase in de evolutiegeschiedenis waarin een gunstig warm klimaat achtergebleven groepen nog wat respijt bood voordat ze zouden worden vervangen door geavanceerde vormen. Bij veel van die groepen, zoals de dinosauriërs en de zeereptielen, was het overigens wat raadselachtig waardoor ze eigenlijk waren uitgestorven — de zoogdieren zouden immers pas na hun heengaan tot bloei komen — maar de pterosauriërs vormden daarop een gelukkige uitzondering: het leek zonneklaar dat ze door de vogels waren weggeconcurreerd.
Onderzoek dat niet in dit beeld paste, werd genegeerd, zoals de conclusie van Ferdinand Broili uit 1927 dat microscopisch onderzoek duidelijk haarzakjes in de huid liet zien[45] en het oordeel van paleoneurologe Tilly Edinger uit hetzelfde jaar dat de hersenen van pterosauriërs meer leken op die van vogels dan reptielen.[46] Wel werd een studie veel geciteerd die het vermogen tot een echte slagvlucht in twijfel trok. Al eind negentiende eeuw waren de luchtvaartpioniers die zweefvliegtuigen probeerden te ontwikkelen geïnspireerd geraakt door Pteranodon. In 1914 verscheen een analyse van het vliegvermogen van dit dier van de hand van Ernest Hanbury Hankin en David Meredith Seares Watson die het afschilderde als een pure zwever.[47]
De pterosauriërrenaissance
In het begin van de jaren zeventig leefde de belangstelling voor pterosauriërs weer op; de paleontoloog Robert Bakker beschreef dit als een "pterosauriërrenaissance".[48] De pterosauriërs voeren mee in het kielzog van de "dinosauriërrenaissance" waarvan dezelfde Bakker de naamgever en medeoorzaak was. Bij de dinosauriërs had Deinonychus al bewezen dat deze groep veel geavanceerder was dan eerst gedacht, en ook de pterosauriërs kenden in 1971 een spectaculaire vondst die hun hele imago verbeterde: Sordes pilosus, waarvan het fossiel onweerlegbaar aantoonde dat de pterosauriërs behaard waren. Toch kwam hierna niet onmiddellijk van wetenschappelijke zijde een krachtige theorievorming op gang. Het was in 1975 wetenschapshistoricus Adrian Desmond die in een populair-wetenschappelijk boek over de theorieën van Bakker, The hot-blooded dinosaurs, ook een hoofdstuk aan de pterosauriërs wijdde, waarin voor het eerst op samenhangende wijze uiteengezet werd waarom ze vermoedelijk warmbloedig waren.[49] Desmond maakte zich daarbij niet helemaal los van de oude denkbeelden: hij zag de grote soorten als zwevers die afhankelijk waren van een stabiel klimaat en zo hulpeloos op de grond dat het een raadsel was hoe ze daar zouden hebben kunnen overleven.
Enkele jaren later besloot Kevian Padian de "Bakker" van de pterosauriërs te worden: in publicaties in 1979,[50] 1980,[51] 1983[52] en 1987[53] verkondigde hij dat pterosauriërs ten onrechte nog steeds geïnterpreteerd werden binnen het verouderde "vleermuismodel" van Von Soemmerring en dat een objectieve analyse aantoonde dat ze niet alleen warmbloedig en behaard waren maar ook een smalle vleugel hadden die niet aan de enkels vastzat, wat de achterpoten bevrijdde voor hun werkelijke rol: er hard op te lopen als teenganger. Pterosauriërs zouden dus gerend hebben als vogels en dat was ook niet zo vreemd omdat ze binnen de Ornithodira ten nauwste aan dinosauriërs verwant zouden zijn geweest.
Padians "Terug naar Seeley"-beweging vond echter geen algemene bijval. Ondertussen waren meer paleontologen zich in het onderwerp gaan specialiseren en die dachten er vaak anders over. De meest invloedrijke daarvan was de Duitser Peter Wellnhofer die in 1978 het eerste standaardwerk over pterosauriërs had laten verschijnen.[54] Hij vond dat pterosauriërs zich op (platte) voeten en vleugels hadden voortbewogen. Ook over het "smalle" vleugelmodel en de "ornithodira-hypothese" ontstonden in de jaren negentig controverses die nog steeds voortduren. Wel veranderde de letterlijke beeldvorming van de pterosauriërs sterk door de illustraties van Gregory S. Paul, die exact de vorm van het skelet volgden en, behalve natuurlijk de vacht, ook de spiergroepen lieten zien.
Al die nieuwe theorieën konden getoetst worden aan een groot aantal nieuwe vondsten. De ontdekking van zeer productieve vindplaatsen, waarvan de belangrijkste de Santanaformatie in Brazilië en de formaties in Liaoning in China zijn, leidde tot een verveelvoudiging van de bekende soorten, waaronder de eerste uit het Trias. Nadat er slechts twee geslachten waren beschreven in de jaren zestig, kwam er vanaf 1970 een constante stroom op gang: tien in de jaren zeventig, twaalf in de jaren tachtig en negen in de jaren negentig. Daarna vond er zelfs een ware explosie van vondsten plaats, waarvan de groei nog niet afgevlakt is:[55] vanaf het jaar 2000 tot midden 2009 zijn 43 geslachten benoemd. Daarna versnelde het tempo zelfs nog. Dat betekent dat van de kleine tweehonderd geslachten het merendeel niet de tijd heeft gehad om bij een ruimer publiek bekend te raken. Er zijn maar een paar populair-wetenschappelijke boeken over het onderwerp verschenen en in de algemenere populaire werken over paleontologie worden steeds dezelfde "klassieke" soorten getoond die we al uit de negentiende eeuw kennen. Een uitzondering vormt Quetzalcoatlus die als "grootste vliegende dier uit de geschiedenis" wordt opgevoerd.
Overigens is ook het aantal technisch-wetenschappelijke boeken over de pterosauriërs gering. Een recent standaardwerk dat alle soorten systematisch behandelt, ontbreekt bijvoorbeeld — een situatie die zich trouwens bij de meeste diergroepen voordoet. Wel worden er meer studies verricht naar de biomechanica van de pterosauriërs die gebruikmaken van computersimulaties die recente inzichten in de aerodynamica toepassen. Ook maakt men sinds begin jaren tachtig vliegende mechanische modellen op ware grootte om die inzichten te toetsen. Het onderzoek naar de groep is zo nog volop in beweging — van de 1828 in 2017 getelde publicaties over pterosauriërs was de helft na 1990 verschenen[56] — en opgravingen worden, nu dergelijke vondsten veel positieve publiciteit opleveren, ook iets ruimer ondersteund door de overheden, hoewel het wereldtotaal de één miljoen dollar per jaar niet overstijgt.
Moderne inzichten over de bouw en levenswijze van de pterosauriërs
De pterosauriërrenaissance heeft vele verbeterde inzichten opgeleverd over de bouw en levenswijze van de pterosauriërs, die aanzienlijk afwijken van hoe nog vrij kort geleden over de groep gedacht werd. Informatie in werken voor 1978 geschreven, populair-wetenschappelijk of wetenschappelijk, is daarmee sterk verouderd geraakt. Veel vraagstukken zijn echter ook tegenwoordig nog niet opgelost; over het geheel genomen toont onze kennis grote hiaten.
Kennisbronnen
Het grootste obstakel voor het begrijpen van de pterosauriërs vormt het feit dat ze er niet meer zijn: hun uitsterven heeft ons beroofd van een goed primair interpretatiemodel. Ook de populaire methode van het phylogenetic bracketing: het in beginsel als algemeen aanwezig voor een groep aannemen van een eigenschap omdat zij zowel voorkomt bij een afstammeling als bij de nauwste verwant in een zijtak, kan hierom meestal niet worden toegepast — nog afgezien van het probleem dat de identiteit van de nauwste verwanten omstreden is. De onzekerheid over de afkomst van de groep, gecombineerd met haar hoge specialisatie, maakt het erg riskant parallellen te trekken met andere groepen. De hele geschiedenis van het pterosauriëronderzoek staat in het teken van een speurtocht naar een fundamenteel referentiekader voor de pterosauriërs, maar dan buiten de groep zelf: de vogel, de vleermuis of "het reptiel". De laatste tijd is duidelijker geworden dat pterosauriërs unieke wezens waren met heel eigen oplossingen voor het probleem van het overleven en dat we die alleen kunnen begrijpen door hun fossielen zo veel mogelijk voor zichzelf te laten spreken.

Afgezien van kleine botfragmenten waarvan er tienduizenden verzameld zijn, vooral uit de Cambridge Greensand, zijn fossielen van pterosauriërs tamelijk zeldzaam: tot nu toe zijn er minder dan drieduizend van gevonden, daarvan slechts enkele honderden complete skeletten. Er is wel gezegd dat ze allemaal in de opbergkasten van een enkel kantoortje van een paleontoloog zouden passen. De reden voor die zeldzaamheid ligt in de slechte conservering die veroorzaakt wordt door hun zeer holle botten. De kans dat een pterosauriërskelet intact fossiliseert, is daarom uiterst gering. Slechts bij zeer gunstige omstandigheden zoals snelle sedimentatie of het vallen in zuurstofloos water, zal zo nu en dan een redelijk compleet dier bewaard blijven. Deze omstandigheden doen zich meestal voor bij kustgebieden of meren. Zo een vindplaats met veel meer en betere fossielen noemen we een Konservat-Lagerstätte. Van alle verzamelde pterosauriërfossielen komt 40% uit Lagerstätten in maar één grotere formatie: de Solnhofener kalksteen in Duitsland uit het late Jura. De belangrijkste andere formaties: de Triadische kalksteen uit Noord-Italië, de Liassische leisteen uit Zuid-Duitsland, en uit het vroege Krijt de Santanaformatie in Brazilië en de Jehol-groep in China, zijn aanzienlijk minder productief. Wel veel fossielen levert de Niobrarakalksteen uit het late Krijt van Noord-Amerika op, ongeveer de helft van het totaal, maar de gemiddelde kwaliteit daarvan is aanzienlijk minder; de meerderheid van de complete skeletten is hierdoor van Pterodactylus en Rhamphorhynchus uit Duitsland.
Dit alles betekent dat de kans groot is dat we een vertekend beeld krijgen van de pterosauriërevolutie. Soorten die in bossen of op droge vlakten leefden, lieten geen fossielen achter; "zeepterosauriërs" en "waterpterosauriërs" zijn oververtegenwoordigd. En dat er nog geen echte Lagerstätten uit het late Krijt zijn gevonden, is misschien de werkelijke oorzaak van het kleine aantal bekende geslachten uit die tijd in plaats van een geleidelijk uitsterven. Aan de andere kant leidde de superieure conservering van een Lagerstätte ertoe dat van het in totaal geringe aantal fossielen wel een groot deel van zeer hoge kwaliteit is. Vrij veel fossielen laten de afdrukken of (omzettings)resten van zachte delen zien, zoals de vlieghuid of vacht. Met de moderne synchrotron-techniek kunnen chemische resten veel beter zichtbaar worden gemaakt wat uitzicht biedt op een aanzienlijke uitbreiding van onze kennis. Meestal zijn ook de betere skeletten wel erg platgedrukt; de kalksteen van de Santanaformatie heeft echter de eigenaardigheid vaak grote nodules, kalkbollen, rond fossielen af te zetten zodat ze driedimensionaal bewaard blijven.
Behalve hun lichamen laten dieren ook soms sporen, uitwerpselen (coprolieten) en eieren achter. Bij pterosauriërs is dit soort overblijfselen nog zeldzamer dan hun botten. Na lange controverses heeft de ichnologie, de sporenkunde, een bepaald zeldzaam type spoor als dat van pterosauriërs erkend en dit bevestigt een bepaalde theorie over hun voortbewegen. Het eerste spoor werd vermoedelijk beschreven in 1954.[57] Wat betreft de oölogie of eierkunde zijn enkele nesten ontdekt. Een mogelijke coproliet werd in 2015 voor het eerst gemeld bij een exemplaar van Rhamphorhynchus.[58]
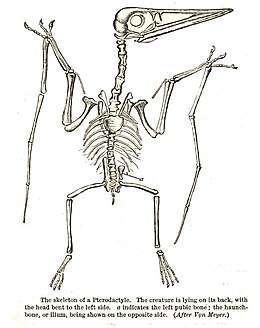
Het skelet
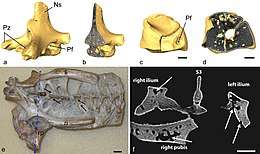
Het skelet van pterosauriërs toont vele eigenaardigheden die uniek zijn voor de groep en door alle soorten gedeeld worden. Deze hangen voornamelijk samen met het vermogen tot vliegen. In sommige opzichten, vooral wat betreft versteviging en gewichtsbesparing, gaan de aanpassingen verder dan de vogels in 160 miljoen jaar evolutie veranderd zijn en dat gegeven is wel gebruikt als argument voor een grote ouderdom van de pterosauriërs. Aan de andere kant tonen de vogels soms extremere modificaties, vooral wat betreft de reductie van de staart en de vergroeiing van schedeldelen en het bekken. Een opvallend kenmerk van de botten van pterosauriërs is hun uiterst lichte constructie. Hoewel het overdreven is de beenwanden als "dun als papier" te omschrijven zoals vaak gebeurt, zijn ze met een typische dikte van één à drie millimeter bij de grotere soorten niet bepaald zwaargebouwd. Vaak is sprake van pneumatisering: het doortrokken zijn met luchtholten die via pneumatische foramina in verbinding staan met luchtzakken, waarlangs ook het vocht dat anders de botten zou vullen, wordt afgevoerd. De lichte buisconstructie die zo ontstaat is wel heel stevig en bijzonder efficiënt. De holle botten zijn op kritieke punten intern verstevigd door een honingraatstructuur of dunne botspanten die werken als stutten of schoren om een fataal buigmoment ("knakken") te voorkomen. De gewichtsbesparing had niet ten doel de totale beenmassa te verminderen — een pterosauriër had ongeveer evenveel bot als een ander gewerveld landdier van dezelfde grootte — maar om voor hetzelfde gewicht de doorsnede van de botten te vergroten zodat ze zwaarder belast konden worden, zowel voor het vliegen als door de hefboomwerking van een grotere schedel.
Hoewel skeletten van de pterosauriërs in het algemeen onderling niet heel veel verschillen, vertonen op sommige punten deelgroepen echter toch aanzienlijke veranderingen; een belangrijke cluster daarvan kenmerkt de Pterodactyloidea. Het skelet wordt traditioneel onderverdeeld in de schedel, een samenstelling van vele beenderen, die zeer informatief kan zijn over afkomst en levenswijze, en de delen achter de schedel, de postcrania.
Postcrania

De romp van de pterosauriërs is sterk gespecialiseerd in het vliegen. Hij is erg kort en heeft een aerodynamische druppelvorm: de voorkant is breed en afgerond en loopt naar achteren haast spits toe zodat de ruimte voor de buikholte zeer beperkt is, de contouren waarvan exact bekend zijn omdat die van onderen begrensd wordt door (zes tot acht) buikribben of gastralia, die als huidverbeningen overigens niet tot het eigenlijke skelet behoren. De brede voorkant draagt een krachtige schoudergordel. De sleutelbeenderen ontbreken of zijn volledig vergroeid met het sternum. Het schouderblad (scapula) is groot, ligt bijna horizontaal en is van voren en onderen verbonden aan een al even stevig ravenbeksbeen (coracoïde); samen vormen ze een os scapulocoracoideum. Op hun verbindingspunt ligt het zijwaarts, naar achteren en iets naar boven gerichte schoudergewricht, met een zadelvormig oppervlak, dat tijdens de vlucht de krachten die de vleugelslag opwekt, moet dragen. Het ravenbeksbeen voert die schuin naar achteren en beneden af naar een borstbeen (sternum) waaraan het verankerd is in een speciaal gewricht. De bevestigingspunten van beide voorste ledematen liggen niet symmetrisch naast elkaar maar achter elkaar, zodat de ravenbeksbeenderen van voren bezien een V-vormige punt vormen. Het borstbeen vormt een plaat die bij de meeste soorten van voren de volle breedte van de borstkas bedekt en via zes of zeven sternale ribben losjes aan de wervelribben verbonden is, die zelf met bovenliggende dubbele gewrichten aan hun wervels scharnieren. Het schouderblad is van boven verankerd aan de ruggengraat, elf tot achttien wervels lang (zonder sacrale wervels), waarvan bij veel soorten de voorste drie tot zeven[59] in meer of mindere mate vergroeid zijn en zo een onbuigzaam notarium vormen. Bij de afgeleide Pterodactyloidea vormen daarin de bovenkanten van de doornuitsteeksels een doorlopende hogergelegen supraneurale plaat met zijdelingse gewrichtsfacetten voor de schouderbladen. Bij elkaar levert het een stijve constructie op, een gesloten beenring, die zonder pezen of kapsels te beschadigen de sterke en langdurige belastingen kan opvangen die het vliegen veroorzaakt en voorkomt dat daardoor de borstkas ingedrukt wordt.
Bij de vogels draagt het borstbeen over de volle lengte een erg hoge kam voor de bevestiging van krachtige borstspieren. Het borstbeen van de pterosauriërs heeft een veel lagere kam maar die loopt over in een ver naar voren uitstekende cristospina of doornkam, aan de basis waarvan de ravensbeksbeengewrichten liggen en die mede de drager is van de twee belangrijkste vliegspieren: zowel de musculus pectoralis, de spier die de vleugel doet neerslaan, als de musculus supracoracoideus die de opslag bewerkstelligt. Het forse opperarmbeen, ondanks zijn korte bouw vaak het in volume grootste bot van het lichaam, is volgens de traditionele interpretatie via een pees die rond een uitsteeksel loopt aan het ravenbeksbeen dat als een katrol werkt aan de musculus supracoracoideus verbonden zodat die ondanks zijn onderliggende positie toch een kracht naar boven kan uitoefenen. Het bestaan van een dergelijk mechanisme is onlangs ontkend door Bennett, die denkt dat alleen de rugspieren de arm hieven.[60] Ook de aan het schouderblad verbonden musculus deltoideus trekt de arm omhoog, terwijl de aan de rug verbonden musculus latissimus dorsi de vleugel naar achteren kan trekken. Al die vliegspieren tezamen vertegenwoordigen het merendeel van de massa van de romp.
Het opperarmbeen is via de elleboog verbonden met de twee beenderen van de onderarm: het spaakbeen en de ellepijp. Die zijn altijd langer dan de bovenarm. Het elleboogsgewricht staat maar beweging toe in één vlak. Als de elleboog wordt gebogen en de vleugel ingeklapt, doet een uitsteeksel van het opperarmbeen het aan de voorzijde gelegen onderarmbeen, het spaakbeen, langs de ellepijp naar voren schuiven en dit duwt via de pols weer de middenhandsbeenderen naar achteren. Zo ontstaat er een automatisch opvouwmechanisme voor de vleugel. Overigens vindt de hoofdbeweging daarvan plaats in de vleugelvinger die vaak een boog kan maken tot 170°: de pols zelf staat maar een beperkte beweging zijwaarts toe en is op en neer nog onbeweeglijker. De twee polsbeenderen, carpalia, die aan de onderarm grenzen, zijn bij afgeleide soorten vergroeid tot een enkelvoudig syncarpale en de vier die de hand raken gereduceerd tot twee, waarvan de grootste, zelf ook een syncarpale vergroeiing van drie carpalia, het vierde middenhandsbeen ondersteunt en de kleinere voorste verbonden is aan een bot dat uniek is voor de Pterosauria: het os pteroides, een dun stiftvormig bot dat naar de romp wijst en vermoedelijk het propatagium ondersteunde, een voorste vlieghuid die van de pols naar de nek liep. Tussen beide syncarpalia is een zekere beweging mogelijk, wat volgens sommige onderzoekers de stijfheid van de hoofdgewrichten compenseert.
Van de middenhandsbeenderen, die bij de Pterodactyloidea langer zijn dan het opperarmbeen, ontbreekt het vijfde; de drie voorste zijn smal en lang en lopen parallel, zonder te vergroeien; ze monden uit in drie korte vingers met respectievelijk één, twee en drie kootjes die eindigen in scherpe klauwen die naar binnen wijzen; de "handpalm" is naar voren gericht. Bij de Pterodactyloidea zijn deze relatief klein. Het veel zwaardere vierde middenhandsbeen draagt de vleugelvinger van vier sterk verlengde kootjes die meestal geleidelijk in grootte afnemen en het merendeel van de spanwijdte van de vleugel uitmaken. Het eerste kootje is met een enorm scharniergewricht aan zijn middenhandsbeen verbonden waarvan het cilindervormige uiteinde de "pin" vormt. Dit gewricht staat slechts een beweging in één vlak toe: het kootje kan als de vleugel zijwaarts uitgeklapt is van voren naar achteren de grootste rotatie van alle zeven vleugeldelen uitvoeren. De kootjes zijn met zadelgewrichten aan elkaar verbonden waarmee de drie uiterste juist een beweging van boven naar beneden kunnen uitvoeren; hun mate van beweeglijkheid is omstreden. Van boven bekeken is de vinger licht naar achteren gekromd — met vaak een extra gekromd uiterste kootje wat het zweefvermogen en de wendbaarheid verbetert[61] — en van voren bezien ook bij de maximale strekking sterk naar beneden gebogen. Reconstructies die het vleugeluiteinde kaarsrecht tonen, zijn dus niet correct. Volgens de traditionele interpretatie scharniert de vleugelvinger zijwaarts ten opzichte van het vierde middenhandsbeen dat dan niet in hetzelfde vlak ligt als de andere vingers. Volgens Bennett echter is dit ten opzichte van de pols een kwartslag naar achteren om zijn as geroteerd, zodat de hele handpalm naar voren wijst en het opvouwen van de vinger overeenkomt met de normale strekking en het naar voren klappen met de buiging.
Als het dier lopend op de middenhandsbeenderen steunt, bevindt de onderarm zich in een toestand van supinatie: het spaakbeen is zo rond de ellepijp gedraaid dat de "handpalmen" naar binnen gekeerd zijn en de vleugelvinger tegen de achterkant van de onderarm opgeklapt is en de drie uiterste kootjes over de rug naar binnen krommen. De drie kleinere klauwen rusten dan zijwaarts schuin naar achteren op de grond, niet naar voren gericht zoals vele illustraties foutief tonen. Pronatie, het naar achteren draaien van de palmen, is vermoedelijk onmogelijk.

De bekkengordel is veel zwakker gebouwd dan de schoudergordel en wordt intern verbonden door de, drie tot tien, sacrale wervels die aan iedere zijde vergroeid zijn met een naar voren uitstekend darmbeen (os ilium), de drager van de voornaamste beenspieren. Dit is lang en plat en van onderen verbonden aan een zitbeen (os ischii) en een schaambeen (os pubis), die bij meer afgeleide soorten meestal vergroeid zijn tot een ischiopubicum, waarvan de geringe hoogte verantwoordelijk is voor het taps toelopen van de romp. Voor beide ossa pubis, maar er niet mee vergroeid, steken schuin naar beneden en voren de smalle prepubes uit die aan hun verbrede uiteinde verbonden zijn. Deze ondersteunen de ingewanden en spelen misschien een rol bij de ademhaling.[62] Deze botten zijn vermoedelijk een nieuwvorming bij de pterosauriërs en niet homoloog aan de prepubes van andere groepen, die niets anders zijn dan uitgroeisels van de ossa pubis.
Tussen het ischiopubicum en het os ilium bevindt zich een schotelvormig niet-geperforeerd heupgewricht dat zijwaarts en schuin omhoog gericht is. Daarin past weer de bolvormige kop van het dijbeen. Die maakt maar een beperkte hoek, 10 tot 45°, met de dijbeenschacht, zodat de achterpoten niet helemaal recht onder het lichaam geplaatst kunnen worden, het meest nog bij de basale vormen. De kop staat de benen verder wel een grote bewegingsvrijheid toe. Het dijbeen is slank gebouwd en vrij recht. Het uiteinde, de trochlea, is via een sterk gebogen kniegewricht, dat enigszins naar buiten gedraaid is, verbonden met het scheenbeen en kuitbeen. Dat laatste is vrij dun en bij de meest afgeleide soorten zelfs helemaal ontbrekend. Het staat het onderbeen, dat altijd langer is dan het bovenbeen, geen asrotatie toe. De enkel is een simpel scharniervlak dat ook weer iets naar buiten gedraaid is, met als gevolg dat als het dijbeen zich in een neutrale zijwaartse positie bevindt men het dier van achteren beziend op de voorkant van de voet kijkt. In een dergelijke houding kan niet gelopen worden en men ziet het dus wel als de normale vluchtpositie. Deze hypothese wordt bevestigd door de speciale vorm van de vijfde teen bij basale pterosauriërs: die heeft twee dunne kootjes waarvan het binnenste extreem lang en het buitenste naar buiten gekromd is — dus naar binnen als de voet naar achteren gedraaid is tijdens de vlucht. Vroeger zag men dit als een aanpassing aan het ondersteboven hangen aan een tak; tegenwoordig denkt men dat tussen deze tenen een vlieghuid gespannen was.
De buitenste vijfde teen, die bij de Pterodactyloidea juist sterk gereduceerd is — volledig of door het verlies van een kootje — wordt ondersteund door een kort middenvoetsbeen. De overige vier middenvoetsbeenderen zijn lang en smal en liggen bovenaan strak tegen elkaar, net als bij de dinosauriërs. Maar ze zijn niet vergroeid en lopen onderaan vrij wijd uit; daarbij blijkt uit het enkelgewricht dat de voet plat op de grond stond. Pterosauriërs waren dus zoolgangers, geen teengangers. De voet had overigens wel een zeer grote bewegingsvrijheid naar achteren. De eerste tot en met vierde teen hebben respectievelijk twee, drie, vier en vijf kootjes; de uiterste daarvan zijn kromme scherpe klauwen, die aanzienlijk verlengd werden door een hoornschacht.
Bij de basale pterosauriërs is de staart erg lang tot wel veertig wervels, een uitzondering is Anurognathus met elf. De basis is beweeglijk, vooral in het horizontale vlak, maar de meer naar achter gelegen wervels zijn aan beide kanten voorzien van zeer lange scherpe uitsteeksels in de lengterichting, vele wervels lang zodat de middelste omgeven worden door een bundel van wel veertig beendraden, die de staart verstijven zodat hij als een vluchtstabilisator gebruikt kan worden. Bij de Pterodactyloidea zijn de staartwervels juist sterk gereduceerd in aantal (vijf tot tweeëntwintig) en grootte en is het staartje dat overblijft meestal erg soepel. Het totale aantal wervels in de wervelkolom werd zo gereduceerd van maximaal zeventig, bij Rhamphorhynchus, tot 33 à 34. De wervels buiten de staart variëren veel minder in aantal: van 25 tot 27. De wervels zijn procoel: hol van voren en bol van achteren.
Het aantal wervels in de nek varieert van zeven tot negen. Dit verschil ontstaat niet door nieuwvorming of reductie van wervels maar door een functieverschuiving tussen die van halswervel en voorste ruggenwervel. Volgens Bennett hebben alle pterosauriërs in wezen negen halswervels. De halswervels zijn lang en stevig en dragen bij de basale vormen ribben. Bij de Pterodactyloidea is de nek zelfs langer dan de romp doordat de individuele halswervels enorm in lengte zijn toegenomen en hebben de wervels onderaan uitsteeksels, exapophysen, om de zijwaartse beweging te beperken; de nekribben ontbreken echter of zijn naadloos vergroeid. De nek is, vooral aan de basis, erg stijf met de grootse bewegingsvrijheid naar beneden; de S-vorm van vogels kan niet worden aangenomen. De schedel kan echter alle kanten op bewogen worden.
Schedel

De schedel van de pterosauriërs is vrij groot — soms langer dan nek en romp samen — en lichtgebouwd. De meest basale soorten hadden nog een tamelijk standaard cranium maar bij meer afgeleide vormen trad een opmerkelijke transformatie op waarbij de voorste botten: de premaxillae, maxillae en voorhoofdsbeenderen, als het ware naar voren werden uitgerekt. Niet alleen ontstond zo een langgerekte snuit maar het verlengingsproces werkte ook in omgekeerde richting zodat het wandbeen, het os squamosum en het os quadratum naar de achterkant van de schedel werden gedrongen. De achterste schedelopeningen, twee per zijde, die alle Diapsida oorspronkelijk bezitten, werden hierdoor echter niet gesloten.
Bij deze soorten is de schedel van achteren breed en overdwars in doorsnee driehoekig zodat de grote oogkassen iets naar voren en boven gericht zijn; dit leverde een zekere mate van stereoscopisch zicht op. De ogen worden ondersteund door een cirkel van botjes, de scleraalring. Voor de oogkassen bevindt zich een opening, de fenestra praeorbitalis, dat bij de Pterodactyloidea met de hooggeplaatste neusgaten zou vergroeien; bij dezen zijn ook de interne neusopeningen in het verhemelte tot één gat versmolten. Bij de Lonchognatha steekt de schedel sterk achter het kaakgewricht uit, dat bij de Pterodactyloidea als een schroefgewricht werkt: bij het openen worden de onderkaken uit elkaar gedrongen, wat wel wordt gezien als een aanpassing aan het opslokken van hele vissen in een keelzak. De onderkaken zijn bij de Lonchognatha lang en dun en vooraan samengegroeid. Bij meer afgeleide groepen is zo soms een derde van kaaklengte versmolten tot smalle massieve plaat.

De in aantal zeer variabele tanden staan met een enkelvoudige wortel in aparte tandkassen en zijn meestal vrij gelijkvormig en kegelvormig. Ze worden steeds vervangen. Eudimorphodon heeft verschillende typen gekartelde tanden en ook nog, als zeer basaal kenmerk dat vaak als argument tegen de ornithodirahypothese gebruikt wordt, tanden in het verhemelte. Sommige soorten hebben een groot aantal kleine tanden; andere juist minder maar wel van enorme lengte zodat ze ver buiten de schedel uitsteken als aanpassing om grote vissen vast te houden. Enkele soorten, zoals Pterodaustro, hebben honderden tanden die gemodificeerd zijn tot een zeef waarmee kreeftjes uit het water gefilterd kunnen worden. Bij de Pterodactyloidea treedt vaak een reductie op: de tanden zijn beperkt tot het voorste stuk van de kaken of ontbreken zelfs geheel. Vermoedelijk waren bij veel soorten de voorkanten van de kaken bedekt met een hoornsnavel of rhamphotheca.
Bij de basale soorten ligt de verbinding met de halswervels, de condylus occipitalis op het achterhoofd, vrij hoog zodat de schedel in het verlengde van de nek staat; bij de Pterodactyloidea ligt die meer onderaan de schedel zodat die een sterkere hoek maakt; zo konden deze soorten ondanks hun meer opgerichte houding bij het lopen toch rechtuit blijven kijken; tijdens de vlucht hielden ze daartoe de nek meer naar boven gekromd.
Bij bijna alle pterosauriërgroepen hebben veel soorten kammen aan de schedel: soms op de snuit, soms achteraan; ook de onderkaak draagt van voren soms een kam. De kammen kunnen van enorme grootte zijn en bizar van vorm. Soms zijn ze verlengd door flappen kraakbeen. De functie ervan is omstreden. De kammen, de voorste uitgroeiingen van het dentarium of het praemaxillare, de achterste van het wandbeen, staan altijd in de lengterichting; die vooraan zijn wel verklaard als aanpassingen voor het stabieler doorklieven van het water tijdens het grijpen van vissen. De achterste schedelkammen zijn geduid als stuurvlakken, dragers van huidzeilen voor extra voortstuwing of als contragewicht voor de lange snuit. Uit moderne simulaties blijkt echter dat ze meestal zo gevormd en geplaatst zijn dat ze juist zo min mogelijk aerodynamisch effect hebben. De meest populaire verklaring is tegenwoordig dan ook dat ze dienden als signalen voor communicatie binnen de soort: als dreiging, geslacht- en leeftijdsonderscheid en als schoonheidskenmerk voor seksuele selectie. Er zijn aanwijzingen bij Pteranodon en Darwinopterus dat de mannetjes de grotere kammen hadden.[63]
Weke delen
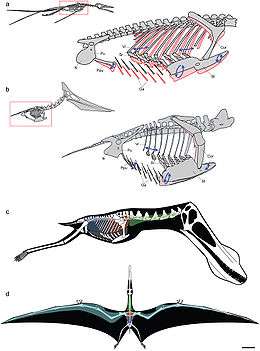
Van de verdere organen naast het skelet, is niet veel bekend. Eén fossiel van Pteranodon toont afdrukken van de luchtpijp, een van Rhamphorhynchus afgietsels van wat darmen. Men neemt aan dat de pterosauriërs net als alle Amniota over huid, bloed, hart, spijsverteringskanaal en gepaarde longen en nieren beschikten. Als Archosauromorpha hadden ze wellicht een cloaca die boven het os ischii uitmondde. Van de spieren kan plaats en sterkte enigszins afgeleid worden van hun aanhechtingspunten op de botten. De brede borstkas bood veel ruimte voor een groot, vierkamerig, hart dat nodig was om de forse vliegspieren te voeden tijdens hun zware inspanningen.
Aan de pneumatische foramina, de openingen in de holle botten, kan men zien dat de longen, al bij de meest basale bekende soorten,[64] verbonden waren met luchtzakken.[65] Die bevonden zich in het achterlijf, langs de wervelkolom op rug en nek en achter de armen. Die zakken maakten het dier wat lichter dan het volume zou doen vermoeden. Behalve voor het ontluchten van de botholten dienden ze om een zeer afwijkend en superieur ademhalingssysteem mogelijk te maken. Bij een luchtzaksysteem, zoals ook de Saurischia (waaronder de vogels) kennen, werken de longen niet meer als een zich met lucht vullende balg met een opening die zowel als in- als uitgang dient. Ze blijven juist constant in volume en hebben een aparte van kleppen voorziene ingang en uitgang zodat de luchtstroom steeds in één richting gaat in plaats van telkens om te keren. Hierdoor is een veel efficiëntere zuurstofopneming mogelijk. Niet de longen zelf drijven de lucht rond, maar afwisselend opblazende luchtzakken vóór en achter de longen, die aangedreven worden door de buikribben, de achterste ribben en bij de pterosauriërs wellicht de beweeglijke prepubes. Bij het inademen vullen zich de achterste luchtzakken en trekken zo de lucht een longhelft in langs de naar binnen openende klep van de ingang; bij het uitademen stoten die luchtzakken de lucht naar voren door de andere longhelft langs de naar buiten openende klep van de uitgang, geholpen door de voorste luchtzakken die zich dan vullen — dit sluit tegelijkertijd de klep van de ingang zodat de luchtstroom niet omkeert.


De allerbeste fossielen laten afdrukken of resten — en een enkele keer zelfs de versteende weefsels — zien van een van de meest relevante onderdelen: de vlieghuid, waarvan de structuur hierdoor vrij exact bekend is. De vleugel werd voornamelijk gevormd door een cheiropatagium, een membraan achter de vleugelvinger. Hoewel nog vaak de vergelijking gemaakt wordt met de vleugels van vleermuizen, gaat deze eigenlijk niet op en ook een vaak gebruikte omschrijving als "leerachtig" is volstrekt onjuist. Niet alleen wordt het membraan bij vleermuizen door meerdere vingers ondersteund, het is ook geheel anders van aard. Bij vleermuizen wordt de vleugel op spanning gebracht door de vleugelhuid te rekken. Bij pterosauriërs wordt de noodzakelijke stijfheid en veerkracht voornamelijk verkregen door een groot aantal dunne (0,1 mm) en kaarsrechte verdikte vezels van bindweefsel (collageen) die in smalle stroken, min of meer parallel, van de vinger schuin naar de achterste vleugelrand lopen en niet uitgerekt kunnen worden. De vezels waren stijver nabij het uiteinde van de vleugels en vrij soepel bij de romp. De eerste zone wordt wel het actinopatagium genoemd en het tweede het tenopatagium. Dit unieke systeem heeft dus meer de eigenschappen van een papieren waaier, zij het dat deze ribbels niet uiteenlopen. De spanning en welving worden bepaald door verbindende spiertjes tussen de ribbels, bestaande uit dwarsgestreept spierweefsel onder bewuste controle van het centraal zenuwstelsel, zodat het vleugelprofiel naar behoeven kan worden ingesteld. Misschien dat dit mede werd geregeld door de luchtzakken in de vleugel; pezen in de membranen ontbreken echter. Bij sommige fossielen kunnen drie lagen worden vastgesteld.[66] Een studie uit 2016 had een afwijkende conclusie dat de vezels in de huid zelf lagen. Dat zou een weinig rekbaar vlies opgeleverd hebben dat alleen kon inklappen op aangeboren vouwlijnen.[67]
Een groot twistpunt is hoever het membraan zich uitstrekte. Alle afdrukken laten vrij smalle vleugels zien alsof de achterste vleugelrand bevestigd was aan het middel. Velen menen dat het met een bocht toch aan de enkels bevestigd was. Dit zou hebben geleid tot een lagere vleugelbelasting en grotere wendbaarheid, maar ook een lagere snelheid. Verschillende fossielen tonen inderdaad stukjes vlieghuid die aan de enkels verbonden zijn, maar dat zou ook een cruropatagium, een door de afwijkende vijfde tenen ondersteunde membraan tussen de achterpoten, kunnen zijn. Dit zat dan, anders dan het uropatagium van vleermuizen, bij de basale vormen niet aan de staart vast. Vroeger werd wel aangenomen dat de reductie bij de Pterodactyloidea van zowel de vijfde teen en de staart zou wijzen op een gespleten uropatagium, waarvan de inkeping nog door het staartje ondersteund werd. Sommige afdrukken laten echter zien dat een cruropatagium bij deze groep naar de basis van de staart liep; de aerodynamische functie van zo'n gespleten cruropatagium is nog onduidelijk.
Ook de voorkant van de vleugel is omstreden. Er bestaat een theorie, tegenwoordig nog verdedigd door Unwin, dat het pteroïde recht naar voren stak zodat het propatagium als een verstelbare vleugelneusklep zou kunnen functioneren en het draagvermogen bij lage snelheden verhoogd werd. Fossielen tonen het pteroïde echter altijd naar het lichaam gekeerd en het is ook onduidelijk hoe het in een voorwaartse positie stabiel gehouden zou kunnen worden.[68]
Verdere vlieghuiden zijn wel aangenomen tussen de drie korte vingers — als stuurvlak — en tussen de tenen; bij de laatste zouden die ook kunnen dienen als een zwemvlies. De fossiele afdrukken zijn echter niet erg duidelijk op dit punt. De basale soorten hebben een huidflap achteraan de staart; deze is asymmetrisch en daarom neemt men tegenwoordig weer aan dat Marsh gelijk had en het functioneerde als een verticaal richtingsroer.

Bij de pterosauriërs is althans de onderkant van de voeten beschubd; de beste afdrukken en sporen laten vrij kleine, ruitvormige, en niet-overlappende schubben zien. Het grootste deel van het lichaam en, zoals recente vondsten tonen, ook de vlieghuid was onbeschubd en bedekt door een dunne opperhuid met een dichte vacht van vrij korte (10 mm) en soepele haren of "pycnofibers". Deze haren waren vermoedelijk niet homoloog aan die van de zoogdieren: dus geen kenmerk geërfd van een gemeenschappelijke voorouder maar een geval van convergente evolutie. Bij afdrukken zijn de haarzakjes duidelijk te zien en in 2003 toonde onderzoek aan dat Goldfuß toch gelijk had gehad en ook de haren zelf vaag zichtbaar waren;[69] bij fossielen, zoals die van Sordes en verschillende vondsten uit China (Beipiaopterus, Jeholopterus, Ningchengopterus), die omzettingsresten tonen, zijn de contouren van de haren vaak perfect aangeduid. De vacht op kop en nek vormde soms manen. In 2019 meldde een studie dat twee exemplaren van de Anurognathidae waren gevonden waarbij de haren aan hun uiteinden gespleten waren zodat een soort borstels ontstonden. Dit is een aanwijzing dat de haren homoloog zijn aan de veren van dinosauriërs waarvan sommige vroege typen ook zo'n borstelvorm hebben.[70]
Over de kleur van de pterosauriërs hebben we slechts indirecte aanwijzingen; we zijn (nog) niet in staat het haarpigment of de irisering vanuit de fossiele haarresten te bepalen. Welke ons bekend zijn, tonen ook geen zichtbaar patroon. In de 19e eeuw, toen het vleermuismodel dominant was, lieten de illustraties meestal een bruine kleur zien; het reptielmodel van het midden van de 20e eeuw prefereerde juist een groene kleur. Beide zouden goede schutkleuren geweest zijn voor de kleinere bosbewonende soorten. Het vogelmodel van de late 20e eeuw gaf de pterosauriërs de typische tegenschaduwing (zwart van boven, wit van onder) van zeevogels. Ook werd het populair pterosauriërs, die als reptielen over een uitstekend vermogen om kleuren te zien zouden hebben kunnen beschikken, te voorzien van allerlei bonte signaalkleuren, waarbij de schedelkammen voor de hand liggende plaatsen waren voor een kleuraccent. Bakker heeft opgemerkt dat soorten als Pterodaustro die van oranje kreeftjes leefden, net als flamingo's door het opgenomen pigment roze gekleurd hadden kunnen zijn.
Stofwisseling
Pterosauriërs waren actieve vliegers en voor dieren van meer dan een paar gram gewicht is vliegen een activiteit die veel inspanning vereist; hierdoor hadden ze veel zuurstof nodig, ze waren dus tachyaeroob. Hun efficiënte long-luchtzaksysteem garandeerde een permanente aanvoer van verse lucht. Die zuurstof diende ter verbranding van brandstof waarvan het hoge verbruik dus een snelle stofwisseling inhield. Door de combinatie van hoge spieractiviteit en een isolerende vacht waren ze tijdens het vliegen endotherm: ze hadden een hoge lichaamstemperatuur (warmbloedigheid) die door de eigen verbranding werd opgewekt. Diezelfde vacht maakte het moeilijk om exotherm te zijn, een dier dat zijn warmte van buiten haalt: hij isoleerde immers ook van de belangrijkste warmtebron: de zon.
Huidige zoogdieren en vogels controleren hun lichaamstemperatuur hormonaal en neuraal binnen nauwe grenzen door de huiddoorbloeding en verdamping (over de longen of van zweet) te variëren. Hun rustmetabolisme — de warmte die de mitochondriën in de cel produceren — blijft daarbij vrij constant ongeacht hun activiteitsniveau. Hierdoor is weliswaar de temperatuur altijd optimaal voor een goede enzymfunctie, maar dit systeem, tachymetabolisme, is ook erg verspillend: ze gebruiken wel tien maal zoveel energie als een poikilotherm, zijn temperatuur wisselend latend, reptiel. Toch is het waarschijnlijk dat de meeste pterosauriërs het bezaten en ook rustend endotherm waren. Voor een vliegend dier biedt het een duidelijk voordeel onmiddellijk zijn volle vliegvermogen te hebben. Ook de vacht is daartoe een aanwijzing: hij kan nauwelijks een andere functie hebben gehad dan het, ook in rusttoestand, handhaven van een temperatuurverschil met de buitenwereld.
Alleen voor de allergrootste soorten, levend in een heet klimaat, is het denkbaar dat ze een systeem gebruikten dat gigantothermie heet: de lichaamstemperatuur controleren door massa. Dat ze daarbij iets afkoelden was bij deze reuzenvormen geen groot probleem: omdat ze in staat waren zonder veel inspanning te blijven zweven, konden ze door middel van een korte maar extra krachtige anaerobe verbranding snel het luchtruim kiezen zonder zich eerst op te warmen. Alleen de ondergrens van de temperatuur kon zo vrijgelaten worden: juist bij grote soorten dreigde oververhitting. Het luchtzaksysteem maakte dan een goede afkoeling mogelijk. Ook deze vormen konden het zich echter niet veroorloven geen tachyaerobe capaciteit te hebben: die was weer nodig om bij ongunstige weersomstandigheden de zweefvlucht te verlengen en zo snel mogelijk weer in staat te zijn tot een anaerobe prestatie.
Voedsel
Pterosauriërs stamden af van roofdieren en zelf waren ze dat ook, zoals hun kleine darmstelsel al aangeeft — hoewel soms gespeculeerd is dat de Tapejaridae met hun hoge en relatief korte schedel zaadeters waren. Sommige kleine bosbewonende soorten, zoals Anurognathus, waren vermoedelijk insecteneters. De lange, soms tandeloze, kop van de Lonchognatha, is traditioneel altijd geïnterpreteerd als een aanwijzing dat de meeste bekende pterosauriërs viseters waren of op andere zeedieren zoals inktvissen joegen. Ze zouden over het wateroppervlak gezweefd hebben om met hun snavel toe te slaan zodra ze een prooi bespeurden. Zelfs werd gedacht dat ze op de wijze van de huidige schaarbekken met hun kaken door de golven schaarden totdat ze op iets eetbaars stuitten. Ook werd aangenomen dat ze in het water doken.

Dit beeld is de laatste tijd wat bijgesteld, vooral wat betreft de grotere soorten. Simulaties tonen aan dat het scharen voor zulke grote dieren biomechanisch uiterst bezwaarlijk is vanwege de te hoge weerstand. Het is nog onduidelijk of de zwaarste vormen van het wateroppervlak hadden kunnen opstijgen. Visvangst vanuit een horizontale vlucht blijft een plausibele methode, zeker bij de basale groepen, maar het recente inzicht dat de Pterodactyloidea zich hadden aangepast aan het lopen op het land, leidde tot de conclusie dat ze daar dus ook hun voedselbronnen hadden. Een eerste hypothese was dat ze geleefd hadden als aaseters zoals de moderne gieren. Hun vermogen om zwevend het landschap af te speuren naar kadavers leek hiermee in overeenstemming. Echter, hun bekken vertonen geen aanpassingen aan het opentrekken van de huid van krengen. Daarom is een andere hypothese populair geworden: het "superooievaar"-model dat stelt dat de Pterodactyloidea leefden als reusachtige steltvogels en aan de waterkant in de modder zochten naar waterdieren.
Lange tijd werd gemeend dat speciale "roofpterosauriërs" die op andere pterosauriërs joegen, niet bekend waren.[71] Aangezien ook de roofvogels in het Mesozoïcum nog niet tot ontwikkeling waren gekomen, hadden de pterosauriërs dan tijdens de vlucht geen belangrijke natuurlijke vijanden — tenzij ze uit de lucht geplukt werden door een theropode. In 2009 werd zo'n rol echter gesuggereerd voor Darwinopterus.
Het voedsel werd niet gekauwd maar heel of in brokken ingeslikt. Bij Pterodaustro is een spiermaag met maagstenen vastgesteld die vermoedelijk gebruikt werd om de schaaldieren te vermalen waaruit het dieet van deze pterosauriër bestond.
Voortbeweging
Vliegen
Pterosauriërs waren sterk aangepast aan een bepaalde wijze van voortbewegen: het vliegen. Hun vermogen zich op eigen kracht door de lucht te bewegen maakte in beginsel gebruik van dezelfde mechanismen die de huidige vogels toepassen: de arm vormt een vleugel die opgeheven wordt en uitklappend met kracht naar beneden en voren[72] geslagen. Bij deze neergaande slag is het gebogen vleugelprofiel, dat werkt volgens de Wet van Bernoulli, iets naar voren gericht zodat de door de beweging veroorzaakte liftkracht niet alleen als een opwaartse kracht het dier in de lucht houdt maar meteen ook een stuwkracht vertegenwoordigt die de luchtweerstand kan overwinnen en een voorwaartse snelheid kan handhaven. Hetzelfde effect hebben de reactiekrachten opgewekt doordat er lucht verplaatst wordt als het vleugelvlak, horizontaler staand, een hoek maakt met zijn bewegingsrichting. Bij de opwaartse slag wordt de vleugel wat ingeklapt en naar achteren en boven bewogen, waarbij wat het Bernouili-effect betreft in wezen weer dezelfde krachten kunnen optreden.
Bij vogels is de vleugel echter gedeeltelijk uit flexibele veren opgebouwd die een aantal speciaal gunstige eigenschappen hebben. Samen maken de veren een perfect vleugelprofiel mogelijk. Bij de neergaande slag buigen de zwakkere vleugeltippen automatisch iets meer naar voren, zodat de lucht aan de onderkant van de vleugel naar buiten en achteren ontsnapt, wat meer stuwkracht oplevert. Bij kleine vogels met een hoge slagfrequentie is het voordeliger de opwaartse slag neutraal met de luchtstroom mee te laten uitvoeren waarbij de vleugel zich draait en op het eind de slagpennen zich openen om de weerstand te verminderen; datzelfde vermogen kan ook ongunstige tipwervels voorkomen.
Lange tijd is gedacht dat de pterosauriërs op deze punten in het nadeel waren. De nieuwe inzichten over het vezelsysteem waarmee de vleugel op spanning gehouden werd, wijzen er echter op dat de pterosauriërs hun eigen bijzondere voordelen bezaten. Het vleugelprofiel kon ermee niet alleen in een ideale vorm voor gemiddelde doeleinden gebracht worden maar ook gecontroleerd aangepast aan speciale omstandigheden, zodat naar behoefte luchtwervels konden worden opgewekt of vermeden en weerstand of stuwkracht gevarieerd. Uit de afdrukken weten we dat de uiterste vleugeluiteinden recht afgesneden waren, wat de geïnduceerde weerstand vermindert.
Behalve het cheiro- of brachiopatagium, de aan de achterkant van de vierde vinger verbonden vlieghuid, waren er nog andere oppervlakken die door op de luchtstroom in te werken, het vlieggedrag beïnvloedden. Tussen de voorrand van de arm en de hals bevond zich ook een membraan, het propatagium. Dit leverde een extra liftkracht op en verlengde het vleugelprofiel. Volgens sommigen was het profiel zelfs instelbaar doordat het propatagium via het os pteroide op en neer bewogen kon worden, maar deze hypothese is zeer omstreden. Vogels kunnen met hun gevederde duim, de alula, die als een neusschoep werkt, de luchtstroom over de bovenzijde van de vleugel bij een grote invalshoek stabiliseren, zodat het dier ondanks een lage snelheid niet overtrokken raakt. Het is wel verondersteld dat een membraan tussen de kleine eerste drie vingers bij pterosauriërs dezelfde functie gehad kan hebben. Ook tussen de poten bevond zich een membraan, het cruropatagium, waarvan het nog onbekend is hoever het zich bij de verschillende taxa uitstrekte. Bij die soorten die een membraan van voet tot voet hadden, kon de liftkracht een groot deel van het gewicht van de benen dragen; het kon ook dienen als remklep voor een abrupte snelheidsvermindering, bijvoorbeeld voor de landing. Sommige afdrukken laten een membraan tussen de tenen zien: als de voeten tijdens de vlucht een verticale positie innamen, met de eerste, binnenste, teen aan de onderzijde, konden ze als roer werken om het gieren te controleren. Basale soorten hadden daarvoor ook een lange staart met een vaantje aan het uiteinde. Onduidelijk is nog welke functie de schedelkammen hierin hadden. Rollen werd gecontroleerd door de vleugels, iets wat wellicht vergemakkelijkt werd door de wel veronderstelde mogelijkheid de uiterste vingerkootjes bij één vleugel naar binnen in te klappen, zodat het draagvermogen aan een kant verminderde. Bij een gespleten cruropatagium kon ook het uitsteken van slechts één been een rol veroorzaken.
De meestal wat grotere pterosauriërs — fossielen die met zekerheid volwassen vormen vertegenwoordigen, hebben een spanwijdte van veertig centimeter of meer — zouden er veel voordeel aan gehad kunnen hebben ook met de opwaartse slag stuwkracht op te wekken. Dat zou dan meer met het binnenste deel van de vleugel moeten gebeuren omdat de vinger daarbij wat ingetrokken werd. Dat past goed bij de hypothese dat de vleugel mede aan de benen bevestigd was en tot aan de enkels doorliep. Pterosauriërs hadden volgens berekeningen gemiddeld een aanzienlijk lagere vleugelbelasting, gewicht-vleugeloppervlakte-verhouding, dan vogels. De uiteinden van de vleugels waren echter smal: ze hadden een grote vleugelslankheid wat de efficiëntie bij het snellere vliegen verhoogt door de parasitaire weerstand te verlagen. Deze combinatie is tegenwoordig typisch voor vogels die zich gespecialiseerd hebben in het zweven zoals de albatrossen. Tijdens een zweefvlucht maakt een dier gebruik van thermiek, stijgwinden of, tijdens het dynamisch zweven, van snelheidsverschillen tussen luchtlagen boven het aardoppervlak. Een gelijksoortige specialisatie komt overeen met de levenswijze van viseter die aan veel pterosauriërs toegeschreven wordt.

Verschillende soorten lijken inderdaad typische zwevers te zijn geweest. Euornithocheira als Pteranodon en Ornithocheirus hadden extreem lange en slanke vleugels en door hun relatief kleine romp een zeer geringe vleugelbelasting. Toch gaat dit "albatrosmodel" op een fundamenteel punt niet op. Albatrossen kunnen vrij zwaar zijn — de reuzenalbatros kan zestien kilogram bereiken — maar zijn geen bijzonder zwaar gespierde vogels. Door hun spanwijdte en gewicht te beperken, hoeven ze niet te investeren in een spierstelsel dat voor een groter dier nodig zou zijn om tijdens slechtere weersomstandigheden te blijven vliegen. Voor een vorm met de bouw van een albatros is een gemiddeld gewicht van circa tien kilogram of een spanwijdte van vier meter ongeveer het praktische maximum.[73] Pteranodon had echter een spanwijdte van ruim zeven meter en sommige ornithocheiriden overtroffen dit nog met enkele meters. In 1974 publiceerde Cherrie Bramwell een studie[74] naar het vliegvermogen van Pteranodon, waarin een gewicht berekend werd van 16,6 kg met een aandrijvende spiermassa van drie kilogram, die het dier voorstelde als een soort levend zweefvliegtuig met een glijgetal van bijna twintig: het zou zich zonder thermiek nauwelijks in de lucht hebben weten te houden, was zelfs voor dynamisch zweven te licht belast en kon niet duiken of eenvoudig van het zeeoppervlak opstijgen. Het zou extreem afhankelijk zijn geweest van klippen aan zee om in de lucht te komen en de stijgwind te leveren om na het vangen van een zware vis weer het land te bereiken. Een nog grotere ornithocheiride had zo zelfs helemaal niet meer kunnen functioneren. Bramwell dacht echter dat Pteranodon door zijn holle botten geen krachtige spieren gehad zou kunnen hebben. Tegenwoordig wordt juist aangenomen dat de sterkte van het skelet door holle botten toeneemt en dat heeft geleid tot de hypothese dat de Euornithocheira een combinatie toonden die bij huidige dieren niet meer bestaat: van een grote vleugelslankheid met een zware spiermassa, die tot 40% uitmaakte van het totale gewicht — dat dan óók een stuk hoger ingeschat wordt: 25 - 50 kg. Dynamisch zweven met een aanzienlijke snelheid wordt zo ook mogelijk.

Er is echter een groep die nog zwaarder gebouwd was dan de Euornithocheira: de Azhdarchidae. Toen azhdarchiden, zoals Quetzalcoatlus, nog slechts uit fragmenten bekend waren, dacht men dat ze dezelfde proporties als de euornithocheiren hadden, wat een spanwijdte van wel twaalf tot vijftien meter opgeleverd zou hebben. Latere vondsten toonden aan dat die maximaal tien à elf meter bedroeg maar dat de romp en het opperarmbeen relatief erg stevig waren. De evolutie van deze groep lijkt dus een ander pad te zijn ingeslagen: weg van de zweefvlucht en in de richting van een krachtige klappende vlucht. De schattingen van hun gewicht zijn daarmee gestegen van 60 tot wel 250 kg; zelfs in dit laatste geval zou nog maar 25% van de massa uit vliegspieren moeten bestaan.[75] De gedachte dat een organisme van zo'n massa überhaupt zou kunnen opstijgen, heeft echter veel weerstand opgeroepen. Extrapolaties van de gemiddelde prestaties bij vogels leken aan te tonen dat geen enkel dier zwaarder dan twaalf kilogram in een horizontale klappende vlucht zou kunnen vliegen.[76] Nu liet de knobbelzwaan met zijn slechts gemiddelde musculatuur en maximaal gewicht van negentien kilogram al zien dat die grens kennelijk doorbroken kon worden, maar 250, of zelfs 100 kg leek een onzinnig hoge raming. Het bezwaar was vooral dat juist in het begin van de vlucht de weerstand relatief hoog moet liggen om ondanks een lage snelheid voldoende opwaartse kracht op te wekken. Deze geïnduceerde weerstand vormt als het ware een obstakel dat genomen moet worden voordat het snelheidsinterval dat het meest efficiënt is, bereikt kan worden. Men nam daarom aan dat grote pterosauriërs van klippen sprongen, met geheven armen hellingen afliepen of zich lieten optillen door een kopwind van boven de 7 m/s. Hierbij ging men ervan uit dat bij de kleinere soorten, net als bij de vogels, de afzet plaatsvond door met de benen op te springen. Bramwell had al in 1974 overwogen of ook niet de, immers veel krachtiger, vleugels zelf hiervoor gebruikt konden worden, maar dat voor Pteranodon verworpen omdat zij de spierkracht te gering achtte. In 2008 echter nam Michael Habib haar idee weer op en toonde modelmatig aan dat Quetzalcoatlus door zijn enorme grootte in één afzetbeweging met de vleugels kon versnellen tot 50 km/u en zo meteen een efficiënte snelheid bereiken.[77] Doordat bij gelijke proporties het gewicht met de derde macht toeneemt en de spierkracht slechts met het kwadraat, zouden de grootste azhdarchiden daarna geen continue horizontale slagvlucht hebben kunnen volhouden: korte krachtsinspanningen om hoogte te winnen, een prooi te vangen of een betere thermiek of stijgwind op te zoeken zouden afgewisseld moeten worden door langere zweefvluchten. Hiervoor zouden spieren met veel krachtige "Type II"-spiervezels het nuttigst geweest zijn; tijdens het zweven konden die dan herstellen van hun snelle verzuring.[78]
Lopen

Al sinds Seeley waren er twee hoofdmodellen voor het lopen van de pterosauriërs: in het ene renden ze als vogels op twee benen en waren ze snelle teengangers; in het andere schuifelden ze onbeholpen en vrijwel plat over de grond, die ze zo snel mogelijk verlieten om een rotswand of boom op te zoeken.
Tegenwoordig zijn beide modellen grotendeels verlaten. Pterosauriërs waren bijna zeker geen teengangers: hun tenen konden daarvoor niet voldoende omhoog gebogen worden ten opzichte van de middenvoetsbeenderen. Lopen op de achterpoten alleen zou lastig geweest zijn omdat het zwaartepunt door de altijd lichte en vaak korte staart te ver naar voren lag. Na lange strijd is er in 1995[79] overeenstemming bereikt dat een in 1957 door William Lee Stokes beschreven bepaald type spoor,[80] de Pteraichnus- ichnospecies, inderdaad aan pterosauriërs kan worden toegeschreven; een overtuigende aanwijzing daarvoor zijn de drie naar achteren uitwaaierende kleine vingers. Het toont duidelijk aan dat ze zich op vier poten voortbewogen en zoolgangers waren. Het spoor is vrij breed en de voeten en handen werden kennelijk niet op de middenlijn geplaatst, zoals bij vogels wel het geval is.
Toch is ook het tweede model onjuist gebleken. Weliswaar waren veel pterosauriërs goede klimmers door hun scherpe klauwen en beweeglijke dijbenen, maar ook op de grond konden ze goed uit de voeten en de latere Pterodactyloidea waren bekwame viervoetige lopers en zelfs hardlopers.
Voor de vroege pterosauriërs ontbraken lange tijd sporenfossielen en dit is wel aangevoerd als argument dat deze dieren onbekwame lopers waren. Met zo'n afwezigheid moet echter voorzichtig worden omgegaan, aangezien de afwezigheid van bewijs geen bewijs is van afwezigheid. De basale vormen werden vermoedelijk gehinderd doordat er een cruropatagium tussen de benen zat. Aan de andere kant waren die meestal nog zo fors dat de lengte van de onderbenen overeenkwam met die van de gecombineerde onderarmen en middenhandsbeenderen zodat ze met een horizontale rug gelijkmatig vooruit konden stappen. Daarbij staken de bovenarmen in een horizontale positie zijwaarts uit en zwenkten naar voren en achteren. De onderarmen waren recht omlaag gericht. Doordat de middenhandsbeenderen zo kort waren, konden ze de pas nauwelijks verlengen; er werden dus maar korte stapjes genomen. Aangezien het lichaam zich dicht bij grond bevond, zullen ze last hebben gehad van sommige obstakels. De meeste onderzoekers nemen aan dat ze niet konden rennen. Een uitzondering vormden sommige soorten, zoals Dimorphodon en Wukongopteridae, die hun bovenarmen meer verticaal konden houden.[81] In 2020 werden de eerste sporen van basale pterosauriërs gemeld. Die toonden aan dat ze goed konden lopen.[82]
De Pterodactyloidea werden niet gehinderd door hun gespleten cruropatagium. Hun middenhandsbeenderen waren vaak erg lang zodat de rug meer verticaal gehouden werd en veel grotere stappen genomen konden worden vanuit de pols. Dat maakte het wel lastiger de beweging te coördineren met de juist kortere benen. Volgens Unwin werden die daarom in een meer verticale positie gehouden. Deze combinatie zou het zelfs mogelijk gemaakt hebben te rennen. Pterodactyloidea hadden vaak gereduceerde klauwen en konden dan slecht klimmen. Kennelijk rustten ze niet in bomen of op klippen maar op de grond waar ze ook vaak hun voedsel zochten. De meeste sporen zijn van deze groep.
Voortplanting
Tot voor kort waren eieren van pterosauriërs onbekend en er werd zelfs gespeculeerd dat ze levendbarend waren. In 2004 bleek echter dat de slechte conservering de oorzaak van hun zeldzaamheid was — ze waren leerachtig met een heel dun calcietlaagje — toen dat jaar de vondst van drie eieren openbaar werd gemaakt: twee in China van nog onzekere soorten[83] en één in Argentinië van Pterodaustro.[84] De eerste bekendmaking, in juni 2004, betrof een nagenoeg perfect geconserveerd 121 miljoen jaar oud fossiel van een pterosauriërembryo, nog in het ei dat 53 millimeter lang en 41 millimeter breed was. Toch zou het dier bij geboorte een spanwijdte van 27 centimeter hebben gehad, voldoende om meteen te hebben kunnen vliegen.[85] In 2015 werd gemeld dat een al eerder beschreven exemplaar van Kunpengopterus, waar een gefossiliseerd ei aangetroffen was dat het stervende wijfje uitgedreven had, nog een ei in het lichaam had. Dat zou erop wijzen dat pterosauriërs twee werkende eileiders hadden, die vermoedelijk om de beurt een ei legden. Althans meer afgeleide pterosauriërs zouden dus niet in één keer een groot legsel gedeponeerd hebben.[86]
Unwin stelt dat geslachten die in vele soorten van verschillend formaat verdeeld werden, zoals Pterodactylus en Rhamphorhynchus, in feite slechts één soort hadden waarvan de verschillende groeistadia abusievelijk voor aparte soorten zijn aangezien. Van Pterodactylus en Pterodaustro zijn veel jonge dieren bekend uit alle groeistadia en die tonen dat de pterosauriërs, anders dan vogels en vleermuizen, onmiddellijk bij het uitkomen min of meer dezelfde proporties als volwassen dieren hadden en dus in principe geen periode in het nest hoefden door te brengen. De dunne eierschaal wijst ook op een bedekking met aarde of rottende vegetatie om te veel vochtverlies te voorkomen.[87] Of er broedzorg was, wordt daardoor twijfelachtig. Zonder constante voeding door de ouders zouden pterosauriërs niet, zoals vogels, heel snel in grootte kunnen toenemen. Een probleem met deze hypothese is dat ze fibrolamellair bot bezaten wat duidt op een vrij snelle groei. Ze kan wel verklaren waarom er weinig kleine pterosauriërsoorten bekend zijn: de jongeren van de grotere soorten zouden deze niches bezet hebben. Binnen Unwins model was er ook geen inwendig gereguleerde groeistop en bleven ze dus als de omstandigheden gunstig waren soms tot op hoge leeftijd langzaam doorgroeien. Dit patroon bestaat ook bij huidige reptielen en leidt tot een grote variabiliteit in grootte tussen "volwassen" individuen, waarvan sommigen tot ware reuzen uitgegroeid zouden zijn.
Intelligentie
Over de intelligentie van pterosauriërs hebben we alleen indirecte aanwijzingen, waarvan de belangrijkste de hersengrootte is. Doordat de fossielen meestal zo platgedrukt zijn, kan de inhoud van de hersenpan, die een aparte langwerpige doosvormige structuur vormde aan het achtereinde van de schedel, vaak lastig bepaald worden. Van sommige goede fossielen is echter een CAT-scan gemaakt en er zijn zelfs een paar natuurlijke afgietsels van de herseninhoud bewaard gebleven. Het encefalisatiequotiënt van pterosauriërs ligt tussen dat van de moderne koudbloedige reptielen en de moderne vogels in. Ze hadden grote lobi optici en enorme evenwichtingsorganen en bijpassende kwabben, de flocculi. Dit lijkt erop te duiden dat ze hun vlucht uitstekend konden coördineren — de grote flocculi regelden daarbij wellicht de instelling van het vleugelprofiel — maar verder geen al te hoge intellectuele capaciteiten bezaten en hun sociaal gedrag niet erg complex was. Maar door hun hoge stofwisseling en goede zuurstofvoorziening moet hun hersencapaciteit toch belangrijk beter geweest zijn dan bij enig huidig koudbloedig reptiel. Bij de Pterodactyloidea waren de grote hersenen daarbij sterk uitgegroeid. Ook waren de evenwichtsorganen bij deze groep wat omhoog gedraaid als aanpassing aan een meer naar beneden gericht hoofd.
Externe links
- The Pterosaur Database
- DinoData Flying Reptiles
- The Pterosaur Net
- Pterosaur FAQ (gearchiveerd op archive.org)
Bronnen, noten en/of referenties
Bronnen
Referenties
|
| Dit artikel is op 7 oktober 2009 in deze versie opgenomen in de etalage. |