Cladistiek
Cladistiek of cladisme is een analysemethode die gebruikt wordt in de bio-systematiek om de evolutionaire relaties tussen organismen te proberen te bepalen. Evolutionaire verwantschap kan alleen worden aangetoond op grond van synapomorfieën: de gemeenschappelijke eigenschappen van alle leden van een groep, die ze geërfd hebben van de laatste gemeenschappelijke voorouder van die groep.
| Orientatie | Cladogrammen | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Verticaal | 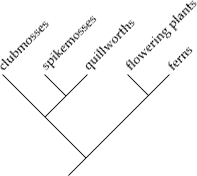 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Horizontaal |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Cladistiek verschilt van andere fylogenetische benaderingen, zoals de evolutionaire systematiek, doordat alleen monofyletische groepen worden opgenomen die alle soorten omvatten die afstammen van een bepaalde voorouder (zogenaamde clades). Afgewezen worden zowel parafyletische groepen – dat zijn groepen die wel alle directe afstammelingen van de laatste gemeenschappelijke voorouder bevatten, maar niet sommige latere afstammelingen – als polyfyletische groepen: groepen die niet alle directe afstammelingen van de voorouder omvatten.
Bij de klassieke systematiek werd voornamelijk gekeken naar de morfologische en anatomische bouw en de overeenkomsten daarin en naar homologieën en overeenkomsten in bouwplan.
De woorden cladistiek en cladisme zijn afgeleid van het Oudgriekse woord κλάδος, klados = "tak". Dit hangt samen met het feit dat de cladistiek zeer veel met stambomen en cladogrammen werkt.
Werkwijze
De constructie van een cladogram verloopt volgens een aantal schijnbaar eenvoudige stappen:[1]
Keuze van de taxa
Kies de taxa of monsters waarvan u de evolutionaire verwantschap wilt onderzoeken. Deze taxa moeten clades zijn als u hoopt tot zinnige resultaten te komen. Soms is het zinnig ook een zustertaxon mee te onderzoeken.
Keuze van de kenmerken
De cladistische methode bestaat uit een analyse van de morfologische en anatomische kenmerken, maar ook fysiologische en genetische kenmerken zoals de volgorde van aminozuren in eiwitten en nucleobasen in DNA.
De te gebruiken kenmerken (eigenschappen) van de organismen worden als eerst bepaald, waarbij zo veel mogelijk relevante kenmerken van de te vergelijken soorten beoordeeld worden of ze ons wel informatie kunnen verschaffen over die verwantschap.
Polariteit van kenmerken
De polariteit van de kenmerken moet bepaald worden, dat wil zeggen: bepaal voor elk taxon of de toestand van een kenmerk 'oorspronkelijk' of 'afgeleid' is. Alle te onderzoeken taxa moeten uniek zijn in zijn kenmerken.
Niet iedere overeenkomst wijst op een nauwe verwantschap: alleen 'afgeleide' of 'nieuwe' kenmerken die bepaalde soorten delen - zogenaamde synapomorfieën - zijn daarvoor een aanwijzing. Om deze synapomorfieën te vinden is het nodig dat alle kenmerken van de te onderzoeken groep soorten systematisch op een rij gezet worden en moet voor elk kenmerk duidelijk zijn wat de oorspronkelijke en wat de afgeleide of geëvolueerde vorm is.
Bij sommige computerprogramma's is deze stap niet absoluut noodzakelijk. Het bepalen van de polariteit van kenmerken kan mede op grond van de kenmerken in "outgroups". Outgroups worden gevormd door een zustergroep of door andere nauwe verwanten van de taxa die het onderwerp van onderzoek vormen. De kenmerken waarin deze outgroup overeenkomt met de te onderzoeken taxa kunnen in principe als 'oorspronkelijk' beschouwd worden.
Synapomorfieën
Om te bepalen welke soorten tot een bepaalde clade behoren zijn alleen gedeelde, 'afgeleide' overeenkomsten tussen soorten, de zogeheten "synapomorfieën", relevant. Dit betekent dat een kenmerk dat gedeeld wordt door twee of meer taxa alleen van enig belang is als het niet in de voorouders van de laatste gemeenschappelijke voorouder aanwezig was maar pas in die voorouder ontstaan is – en dus een afgeleid kenmerk van die voorouder vormt.
Autapomorfieën
Overeenkomsten die later dan de laatste gemeenschappelijke voorouder ontstonden ("autapomorfieën") wijzen niet op een bijzondere verwantschap: vogels en zoogdieren zijn beide warmbloedig maar hun laatste gemeenschappelijke voorouder was dit niet; de overeenkomst is het gevolg van convergente evolutie.
Symplesiomorfieën
Kenmerken die al eerder ontstonden dan de laatste gemeenschappelijke voorouder ("symplesiomorfieën") duiden ook niet op een bijzondere verwantschap: apen en hagedissen hebben beide een staart en zijn nauwer aan elkaar verwant dan aan salamanders; maar de staart wijst daar niet op, want de salamanders hebben die ook als homoloog orgaan.
Groepeer taxa
De taxa worden gegroepeerd aan de hand van synapomorfieën (overeenkomstige afgeleide kenmerken) en niet symplesiomorfieën (oorspronkelijke of "primitieve" kenmerken). Bij de berekening van deze groepen is het vrijwel onvermijdelijk dat bij een grote dataset sommige kenmerken tegenstrijdige informatie opleveren. Deze tegenstrijdigheden worden opgelost met een duidelijk omschreven methode, gewoonlijk parsimonie waarbij het aantal conflicten wordt geminimaliseerd.
Construeer het cladogram
Bouw het cladogram, dat GEEN evolutionaire stamboom is, volgens de volgende regels:
- alle taxa worden geplaatst op eindpunten van het cladogram, nooit bij de knopen (vertakkingspunten).
- iedere knoop van het cladogram moet voorzien zijn van een lijst van synapomorfieën, die gemeenschappelijke kenmerken vormen voor alle taxa boven de knoop (tenzij ket kenmerk in een later stadium is gewijzigd).
- alle synapomorfieën verschijnen slechts een maal in het cladogram tenzij door parallelle evolutie onafhankelijk is ontstaan.
Berekeningen
Bij een klein aantal soorten en weinig eigenschappen, lukt de berekening van een cladogram nog handmatig, bij grotere gegevensverzamelingen wordt dit al gauw te tijdrovend. Tegenwoordig worden cladistische analyses daarom voor een groot deel door computerprogramma's uitgevoerd. Dit wordt in de hand gewerkt doordat sinds de jaren tachtig steeds vaker DNA-sequenties (ook wel "moleculaire data" genoemd) gebruikt worden, wat leidt tot zeer uitgebreide gegevensverzamelingen. De computer berekent die stambomen die het minste aantal "stappen" (veranderingen in vorm) vergen om alle onderzochte soorten in een evolutionair verband te brengen en dus het waarschijnlijkst zijn.
Resultaten
De resultaten van de berekeningen kunnen worden weergegeven in een stamboom of cladogram (of kladogram). Elk cladogram moet altijd bezien worden in het licht van de data die eraan ten grondslag liggen. Een andere dataset voor dezelfde organismen zal allicht leiden tot een ander cladogram. Een voorbeeld daarvan vormt het cladogram voor de bedektzadigen, zoals die er uitziet in het APG IV-systeem voor de "COM"-groep (de Celastrales, Oxalidales en Malpighiales). Op grond van kern-DNA en mitochondriaal DNA krijgt deze "COM"-groep een andere plaats in het systeem dan op grond van het chloroplast DNA.
|
| → benoemde clades zijn omlijnd ← |
Weergave van een cladogram
| lezen van een horizontaal cladogram zonder tijdsas met wortel: LUCA (de hypothetische voorouder van alle hedendaagse organismen) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| knoop Eukaryota benoemd, basale clade boven, kroongroep beneden |
knoop Eukaryota onbenoemd; basale clade beneden | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| takken even lang | takken ongelijk lang en anders verbonden | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Een cladogram omvat een voorouder met alle afstammelingen. Bij onvoldoende kennis is of bij onzekerheden wordt dat weergegeven met een "harkje". Voorbeeld: de vier zustergroepen in de kroongroep Eukaryota.
Er staan alleen namen aan de uiteinden van de takken van een cladogram, de knopen krijgen in een cladogram geen naam, maar de in een taxonomische indeling voorkomende hogere taxa worden soms wel genoemd ter verduidelijking: zie Eukaryota, wat staat voor de hypothetische voorouder van de eukaryoten.
Geschiedenis
Fylogenetische systematiek kwam op na het baanbrekende werk van Charles Darwin. De eerste fylogenetische systemen verschenen tegen het eind van de negentiende eeuw. Cladistiek werd pas in het midden van de twintigste eeuw een aparte discipline. De entomoloog Willi Hennig wordt in het algemeen beschouwd als de "vader" van de cladistiek.
Tot in de jaren zeventig was het ondoenlijk om handmatig de gegevens verder volledig te verwerken. Maar in de jaren tachtig zorgde de voortgeschreden computertechniek ook voor een nieuwe doorbraak binnen de cladistiek. Een computerprogramma bezit tegenwoordig zeer veel rekenkracht en is in staat uit te rekenen welke stamboom het geringste aantal evolutionaire "stappen" (veranderingen) vereist en daarmee de waarschijnlijkste is. Meteen levert dat de synapomorfieën op van alle aftakkingen van de stamboom. Zo'n tak heet een clade (of klade) die bestaat uit de laatste gemeenschappelijke voorouder – de soort die zich van de stam heeft afgesplitst en zo de tak vormde – en al zijn afstammelingen.
De kladistiek werd de kernmethode in de moderne fylogenie, als deel van een algemene verschuiving in de wetenschap in de richting van de kwantitatieve methode. Het subjectieve oordeel van de onderzoeker die eerst moest afgaan op een zeer algemene indruk van de, vaak misleidende, gelijkenis in bouw, hoogstens gecorrigeerd door wat belangrijke synapomorfieën, werd vervangen door een veel objectiever proces dat een exacte uitkomst kon opleveren.
Cladistiek versus Linneaanse classificatie
Een omstreden kwestie is de vraag óf en zo ja in hoeverre de cladistiek uitdrukking behoort te vinden in de taxonomische classificatie. Sommige cladisten willen alleen monofyletische groepen in de classificatie opnemen, maar dat wordt niet door alle taxonomen aanvaard. Andere cladisten menen dat classificatie geen wetenschappelijke activiteit is en dat hun eigen project alleen moet gaan over het toetsen van mogelijke afstammingsverwantschappen zonder zich druk te maken om de formele indeling in allerlei rangen.
Aangezien clades strikt dichotoom zijn is het niet mogelijk alle evolutionaire processen met behulp van clades weer te geven. Netwerkevolutie, waarbij separate clades fuseren, is inherent en natuurlijk polyfyletisch van aard. De clade is een te beperkte topologie om zonder hulpgrepen met deze veel voorkomende processen om te gaan, in plaats van de klassieke stamboom.[6]
Een belangrijke verschil tussen de klassieke Linneaanse classificatie en de cladistiek is dat de eerste een hiërarchische ordening zoekt terwijl de andere alleen de afstammingslijnen aan het licht wil brengen. In de thans meest gangbare benadering die is gebaseerd op min of meer regelmatige mutaties in bepaalde DNA-sequenties is de afstand tussen de claden uitsluitend de weerslag van de tijd die is verlopen sinds de scheiding, zonder de direct waarneembare veranderingen aan de taxa mee te wegen. Veranderingen in gedrag, bouw, fysiologie en ecologie zijn veeleer het gevolg van selectiedruk dan van tijdverloop.
R.K. Brumitt verwoordde de bezwaren als volgt:
|
Linneaanse classificatie zonder parafyletische taxa is logisch ondenkbaar. Ieder monofyletisch genus in een Linneaanse classificatie stamt onvermijdelijk af van iets in een ander genus, die dan dus parafyletisch wordt. Evenzo moet een monotypische familie afstammen van een soort in een geslacht in een andere familie. Als parafyletische taxa niet mogen bestaan, waar komen genera en families dan vandaan? (...) Linneaanse classificatie zonder parafyletische taxa is onzinnig. Hennigs voorstel om parafyletische taxa te elimineren was gebaseerd op de fout niet in te zien dat het verschil tussen de Linneaanse hiërarchie, waarin alle taxa deel uitmaken van het hogere taxon, en een fylogenetische hiërarchie waarin dat niet het geval is en de lagere niveaus niet equivalent zijn aan de hogere. Anders gesteld: alle soorten van een genus zijn het genus, maar alle afstammelingen van een voorouder zijn niet die voorouder. |
Linneaanse en cladistische indelingen vullen elkaar aan. Polyfyletische taxa van hogere rang dan soort[7] behoren ook in een Linneaanse classificatie niet voor te komen en het cladistisch onderzoek levert reeds veel informatie om dit soort onvolkomenheden te herstellen.
Naamgeving van clades
Een minder controversiële vraag is of elke clade die door de analyse ontdekt wordt dan ook werkelijk een naam moet krijgen. Omdat een cladogram zeer veel clades kan bevatten, probeert men een keuze te maken door alleen namen te geven aan relevante relaties tussen interessante soorten. Dit maakt de naamgeving stabieler en geeft wat richting aan de theorievorming. Het vormt ook een subjectief element binnen een methode die poogt juist zo objectief mogelijk te zijn. De naamgeving geschiedt doordat een clade gedefinieerd wordt door óf synapomorfieën als indicaties van afstamming, óf pure afstamming, óf een combinatie van beide. De tweede methode wordt steeds meer gebruikelijk. Definities hebben dan de vorm: "clade K is de groep omvattende de laatste gemeenschappelijke voorouder van soorten X en Y en al zijn afstammelingen" (nodeclade genaamd vanwege het "knooppunt" dat de voorouder vormt) of "clade K is de groep omvattende soort X en alle soorten die nauwer verwant zijn aan soort X dan aan soort Y" (stemclade genaamd omdat een afgesplitste aftakking gedefinieerd wordt).
Hoewel dit niet strikt noodzakelijk is voor het hanteren van de cladistische methode, pleegt men hierbij om principiële of praktische redenen de oude rangonderscheidingen van de taxonomie achterwege te laten, met uitzondering van de geslachts- en soortnamen omdat deze beiden gebruikt worden om een soort mee te identificeren.
Zie ook
| Zie de categorie Cladistics van Wikimedia Commons voor mediabestanden over dit onderwerp. |