VDAC1
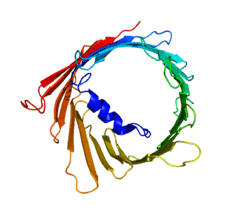


Voltage-dependent anion-selective channel 1 (VDAC-1) is a beta barrel protein that in humans is encoded by the VDAC1 gene located on chromosome 5.[4][5] It forms an ion channel in the outer mitochondrial membrane (OMM) and also the outer cell membrane. In the OMM, it allows ATP to diffuse out of the mitochondria into the cytoplasm. In the cell membrane, it is involved in volume regulation. Within all eukaryotic cells, mitochondria are responsible for synthesis of ATP among other metabolite needed for cell survival. VDAC1 therefore allows for communication between the mitochondrion and the cell mediating the balance between cell metabolism and cell death. Besides metabolic permeation, VDAC1 also acts as a scaffold for proteins such as hexokinase that can in turn regulate metabolism.[6]
This protein is a voltage-dependent anion channel and shares high structural homology with the other VDAC isoforms (VDAC2 and VDAC3), which are involved in the regulation of cell metabolism, mitochondrial apoptosis, and spermatogenesis.[7][8][9][10] Over expression and misregulation of this pore could lead to apoptosis in the cell leading to a variety of diseases within the body. In particular, since VDAC1 is the major calcium ion transport channel, its dysfunction is implicated in cancer, Parkinson's (PD), and Alzheimer's disease.[11][12][13]

Structure
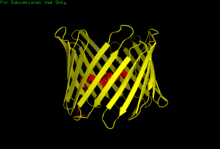
The three VDAC isoforms (VDAC1, VDAC2, and VDAC3) have highly conserved DNA sequences as well as 3D structures forming a wide β-barrel structure, inside of which the alpha helical N-terminal segment resides to partially close the pore.[14] VDAC1's structure was solved by 3 independent labs by x-ray crystallography, Nuclear Magnetic Resonance (NMR) spectroscopy, or a combination of both. Two of these structural studies were used to determine human VDAC1 (hVDAC1) structure while X-ray crystallography was used to solve murine VDAC1 (mVDAC1) structure that differs from hVDAC1 by only two residues.[15][16][17] These determined structures aligned with earlier circular dichorism studies that predicted the presence of alpha helix and β-strand domains.[18]
Structural analysis of mVDAC1's structure showed a barrel-like channel composed of 19 amphipathic β-strands, with the N-terminus and C-terminus both facing towards the inter membrane space of the mitochondrion.[19][20] β-strands are connected via loops and are arranged in an anti-parallel pattern with the exception of β-strands 1 and 19 which are parallel.[17] The pore has a height of 40 Ẳ, spans a distance of 27 Ẳ by 20 Ẳ at the openings and tapers down to 20 Ẳ by 14 Ẳ at the N-terminal α-helix segment in the open state.[21] The closed state conformation has yet to be isolated and determined. Additionally, the N-terminus has an alpha helical segment that is held to the inside wall of the pore by hydrophobic interactions with residues on β-sheets 8-18.[17] This N-terminus can serve as a scaffold for the movement of ions or attachment of proteins. One such example is seen as it is the docking site for HK1 binding.[6] A significant residue to point out is the glutamate located at the 73rd residue on the amino acid chain (E73). This residue is found in VDAC1 and VDAC2 but not VDAC3. The side chain of this charged residue points into the phospholipid bilayer which would normally cause repulsive forces to occur. E73 however, has been implicated in VDAC1 function and interaction.[22]
Function
VDAC1 belongs to the mitochondrial porin family and is expected to share similar biological functions to the other VDAC isoforms.[23] Of the three isoforms, VDAC1 is the main calcium ion transport channel and the most abundantly transcribed.[12][24] VDAC1 is involved in cell metabolism by transporting ATP and other small metabolites across the outer mitochondrial membrane (OMM) allowing regulation of the TCA cycle and, by extension, reactive oxygen species (ROS) production.[11] In yeast cells, ROS accumulates in response to oxidative stress, which results in impaired mitochondrial function and a “petite” phenotype. However, petite yeast cells exhibit a longer lifespan than wild-type cells and indicate a protective function by VDAC1 in similar circumstances, such as aging.[6][24]
Voltage gating
VDAC1 allows for the conductance of molecules into and out of the mitochondrion. Its permeability is dependent on VDAC1's conformational state which is determined by voltage. At low voltage (10mV), the pore is in an "open" state where the channel is weakly anion selective and allows for a greater flux of metabolites. Because of the large pore size, metabolic gating under saturated ATP conditions reveal a transport of 2,000,000 ATP/second and a transport of 10,000 ATP under physiological conditions.[25] At a higher voltage in the positive or negative direction (>30mV), the pore is in a "closed" state and is weakly cation selective allowing for less metabolites to be transported.[17] The flux of metabolites can be seen as negligible.This change in states is mediated by a conformational change in the protein that has yet to be discovered. Since the alpha helical N-terminus segment is located in the center of the pore, it is ideally situated for metabolic gating. This lead researchers to believe that the Alpha helix was a key contributor to determining the conformational states. However, more recent studies have shown the N-terminal is unnecessary for proper voltage gating and therefore suggest the flexible beta barrel as the mechanism of conformational change.[21]
Oligomerization
Atomic Force Microscopy (AFM) revealed the presence of VDAC1 monomers as well as dimers and larger oligomers showcasing the interaction of the pore with itself, however, dimers are more frequent.[26] hVDAC1 in particular has been shown to arrange in parallel dimers leading to increased permeability of the pore.[15] The glutamate located at the 73rd position on VDAC1 has also been shown to play a role in oligomerization when in the presence of calcium.[22] VDACs can also oligomerize to form part of the mitochondrial permeability transition pore (MPTP) and, thus, facilitate cytochrome C release, leading to apoptosis. VDACs have also been observed to interact with pro- or antiapoptotic proteins, such as Bcl-2 family proteins and kinases, and so may contribute to apoptosis independently from the MPTP.[23]
Clinical significance
The voltage dependent anion channels all function in ion and metabolite transport although their physiological roles are different. Because of their role, dysfunction of the channels can lead to various diseases. VDAC1 has been implicated in cancer through its interactions with the antiapoptotic family of proteins, Bcl-2 proteins, particularly Bcl-xl, and Mcl-1, which are overexpressed during cancer. These two Bcl-2 proteins interact with VDAC1 to regulate calcium ion transport across the OMM and, ultimately, ROS production. While high levels of ROS induce cell death, non-lethal levels interfere with signal transduction pathways that can then promote cell proliferation, migration, and invasion in cancer cells.[11] Moreover, VDAC1 overexpression has been associated with increased apoptotic response and anti-cancer drugs and treatment efficacy, further supporting VDAC1 as a therapeutic target for cancer treatment.[11][27]
VDAC1's function in calcium ion transport has also been linked to neurodegenerative diseases. In PD, VDAC1 increases calcium ion levels within the mitochondria, resulting in increased mitochondrial permeability, disrupted mitochondrial membrane potential, elevated ROS production, cell death, and neuronal degeneration.[12] VDAC1 has been shown to interact with Amyloid β (Aβ) leading to increased conductance of the channel and eventually apoptosis of the cell.[13]
Interactions
VDAC1 acts as a scaffold for many proteins as well as allows for the flux of ions and metabolites through interactions within the pore.
A major metabolite that moves through this channel is ATP. A low affinity binding site used for fast transport of this molecule was discovered by the Markov state modeling approach. It was shown that ATP binds to multiple basic residues within the pore sequentially, in essence moving through the channel.[28]
VDAC1 has also been shown to interact with:
See also
References
- 1 2 3 GRCm38: Ensembl release 89: ENSMUSG00000020402 - Ensembl, May 2017
- ↑ "Human PubMed Reference:".
- ↑ "Mouse PubMed Reference:".
- ↑ Blachly-Dyson E, Baldini A, Litt M, McCabe ER, Forte M (March 1994). "Human genes encoding the voltage-dependent anion channel (VDAC) of the outer mitochondrial membrane: mapping and identification of two new isoforms". Genomics. 20 (1): 62–7. doi:10.1006/geno.1994.1127. PMID 7517385.
- ↑ "Entrez Gene: VDAC1 voltage-dependent anion channel 1".
- 1 2 3 4 Reina S, Palermo V, Guarnera A, Guarino F, Messina A, Mazzoni C, De Pinto V (July 2010). "Swapping of the N-terminus of VDAC1 with VDAC3 restores full activity of the channel and confers anti-aging features to the cell". FEBS Letters. 584 (13): 2837–44. doi:10.1016/j.febslet.2010.04.066. PMID 20434446.
- ↑ Subedi KP, Kim JC, Kang M, Son MJ, Kim YS, Woo SH (February 2011). "Voltage-dependent anion channel 2 modulates resting Ca²+ sparks, but not action potential-induced Ca²+ signaling in cardiac myocytes". Cell Calcium. 49 (2): 136–43. doi:10.1016/j.ceca.2010.12.004. PMID 21241999.
- 1 2 Alvira CM, Umesh A, Husted C, Ying L, Hou Y, Lyu SC, Nowak J, Cornfield DN (November 2012). "Voltage-dependent anion channel-2 interaction with nitric oxide synthase enhances pulmonary artery endothelial cell nitric oxide production". American Journal of Respiratory Cell and Molecular Biology. 47 (5): 669–78. doi:10.1165/rcmb.2011-0436OC. PMC 3547107. PMID 22842492.
- ↑ Cheng EH, Sheiko TV, Fisher JK, Craigen WJ, Korsmeyer SJ (July 2003). "VDAC2 inhibits BAK activation and mitochondrial apoptosis". Science. 301 (5632): 513–7. doi:10.1126/science.1083995. PMID 12881569.
- ↑ Li Z, Wang Y, Xue Y, Li X, Cao H, Zheng SJ (February 2012). "Critical role for voltage-dependent anion channel 2 in infectious bursal disease virus-induced apoptosis in host cells via interaction with VP5". Journal of Virology. 86 (3): 1328–38. doi:10.1128/JVI.06104-11. PMC 3264341. PMID 22114330.
- 1 2 3 4 5 Huang H, Shah K, Bradbury NA, Li C, White C (October 2014). "Mcl-1 promotes lung cancer cell migration by directly interacting with VDAC to increase mitochondrial Ca2+ uptake and reactive oxygen species generation". Cell Death & Disease. 5: e1482. doi:10.1038/cddis.2014.419. PMC 4237246. PMID 25341036.
- 1 2 3 Chu Y, Goldman JG, Kelly L, He Y, Waliczek T, Kordower JH (September 2014). "Abnormal alpha-synuclein reduces nigral voltage-dependent anion channel 1 in sporadic and experimental Parkinson's disease". Neurobiology of Disease. 69: 1–14. doi:10.1016/j.nbd.2014.05.003. PMID 24825319.
- 1 2 Smilansky A, Dangoor L, Nakdimon I, Ben-Hail D, Mizrachi D, Shoshan-Barmatz V (December 2015). "The Voltage-dependent Anion Channel 1 Mediates Amyloid β Toxicity and Represents a Potential Target for Alzheimer Disease Therapy". The Journal of Biological Chemistry. 290 (52): 30670–83. doi:10.1074/jbc.M115.691493. PMC 4692199. PMID 26542804.
- ↑ Amodeo GF, Scorciapino MA, Messina A, De Pinto V, Ceccarelli M (2014). "Charged residues distribution modulates selectivity of the open state of human isoforms of the voltage dependent anion-selective channel". PLoS One. 9 (8): e103879. doi:10.1371/journal.pone.0103879. PMC 4146382. PMID 25084457.
- 1 2 Bayrhuber M, Meins T, Habeck M, Becker S, Giller K, Villinger S, Vonrhein C, Griesinger C, Zweckstetter M, Zeth K (October 2008). "Structure of the human voltage-dependent anion channel". Proceedings of the National Academy of Sciences of the United States of America. 105 (40): 15370–5. doi:10.1073/pnas.0808115105. PMC 2557026. PMID 18832158.
- ↑ Hiller S, Garces RG, Malia TJ, Orekhov VY, Colombini M, Wagner G (August 2008). "Solution structure of the integral human membrane protein VDAC-1 in detergent micelles". Science. 321 (5893): 1206–10. doi:10.1126/science.1161302. PMC 2579273. PMID 18755977.
- 1 2 3 4 Ujwal R, Cascio D, Colletier JP, Faham S, Zhang J, Toro L, Ping P, Abramson J (November 2008). "The crystal structure of mouse VDAC1 at 2.3 A resolution reveals mechanistic insights into metabolite gating". Proceedings of the National Academy of Sciences of the United States of America. 105 (46): 17742–7. doi:10.1073/pnas.0809634105. PMC 2584669. PMID 18988731.
- ↑ Shanmugavadivu B, Apell HJ, Meins T, Zeth K, Kleinschmidt JH (April 2007). "Correct folding of the beta-barrel of the human membrane protein VDAC requires a lipid bilayer". Journal of Molecular Biology. 368 (1): 66–78. doi:10.1016/j.jmb.2007.01.066. PMID 17336328.
- ↑ McCommis KS, Baines CP (June 2012). "The role of VDAC in cell death: friend or foe?". Biochimica et Biophysica Acta. 1818 (6): 1444–50. doi:10.1016/j.bbamem.2011.10.025. PMC 3288473. PMID 22062421.
- ↑ Tomasello MF, Guarino F, Reina S, Messina A, De Pinto V (December 2013). "The voltage-dependent anion selective channel 1 (VDAC1) topography in the mitochondrial outer membrane as detected in intact cell". PLoS One. 8 (12): e81522. doi:10.1371/journal.pone.0081522. PMC 3855671. PMID 24324700.
- 1 2 Teijido O, Ujwal R, Hillerdal CO, Kullman L, Rostovtseva TK, Abramson J (March 2012). "Affixing N-terminal α-helix to the wall of the voltage-dependent anion channel does not prevent its voltage gating". The Journal of Biological Chemistry. 287 (14): 11437–45. doi:10.1074/jbc.M111.314229. PMC 3322836. PMID 22275367.
- 1 2 Keinan N, Pahima H, Ben-Hail D, Shoshan-Barmatz V (July 2013). "The role of calcium in VDAC1 oligomerization and mitochondria-mediated apoptosis". Biochimica et Biophysica Acta. 1833 (7): 1745–54. doi:10.1016/j.bbamcr.2013.03.017. PMID 23542128.
- 1 2 Lee MJ, Kim JY, Suk K, Park JH (May 2004). "Identification of the hypoxia-inducible factor 1 alpha-responsive HGTD-P gene as a mediator in the mitochondrial apoptotic pathway". Molecular and Cellular Biology. 24 (9): 3918–27. doi:10.1128/mcb.24.9.3918-3927.2004. PMC 387743. PMID 15082785.
- 1 2 De Pinto V, Guarino F, Guarnera A, Messina A, Reina S, Tomasello FM, Palermo V, Mazzoni C (2010). "Characterization of human VDAC isoforms: a peculiar function for VDAC3?". Biochimica et Biophysica Acta. 1797 (6–7): 1268–75. doi:10.1016/j.bbabio.2010.01.031. PMID 20138821.
- ↑ Rostovtseva, T.; Colombini, M. "VDAC channels mediate and gate the flow of ATP: implications for the regulation of mitochondrial function". Biophysical Journal. 72 (5): 1954–1962. doi:10.1016/s0006-3495(97)78841-6.
- ↑ Hoogenboom BW, Suda K, Engel A, Fotiadis D (July 2007). "The supramolecular assemblies of voltage-dependent anion channels in the native membrane". Journal of Molecular Biology. 370 (2): 246–55. doi:10.1016/j.jmb.2007.04.073. PMID 17524423.
- 1 2 Weisthal S, Keinan N, Ben-Hail D, Arif T, Shoshan-Barmatz V (October 2014). "Ca(2+)-mediated regulation of VDAC1 expression levels is associated with cell death induction". Biochimica et Biophysica Acta. 1843 (10): 2270–81. doi:10.1016/j.bbamcr.2014.03.021. PMID 24704533.
- ↑ Choudhary, Om P; Paz, Aviv; Adelman, Joshua L; Colletier, Jacques-Philippe; Abramson, Jeff; Grabe, Michael. "Structure-guided simulations illuminate the mechanism of ATP transport through VDAC1". Nature Structural & Molecular Biology. 21 (7): 626–632. doi:10.1038/nsmb.2841. PMC 4157756. PMID 24908397.
- 1 2 Weng C, Li Y, Xu D, Shi Y, Tang H (March 2005). "Specific cleavage of Mcl-1 by caspase-3 in tumor necrosis factor-related apoptosis-inducing ligand (TRAIL)-induced apoptosis in Jurkat leukemia T cells". The Journal of Biological Chemistry. 280 (11): 10491–500. doi:10.1074/jbc.M412819200. PMID 15637055.
- 1 2 Shi Y, Chen J, Weng C, Chen R, Zheng Y, Chen Q, Tang H (June 2003). "Identification of the protein-protein contact site and interaction mode of human VDAC1 with Bcl-2 family proteins". Biochemical and Biophysical Research Communications. 305 (4): 989–96. doi:10.1016/s0006-291x(03)00871-4. PMID 12767928.
- ↑ Shimizu S, Konishi A, Kodama T, Tsujimoto Y (March 2000). "BH4 domain of antiapoptotic Bcl-2 family members closes voltage-dependent anion channel and inhibits apoptotic mitochondrial changes and cell death". Proceedings of the National Academy of Sciences of the United States of America. 97 (7): 3100–5. doi:10.1073/pnas.97.7.3100. PMC 16199. PMID 10737788.
- ↑ Shimizu S, Narita M, Tsujimoto Y (June 1999). "Bcl-2 family proteins regulate the release of apoptogenic cytochrome c by the mitochondrial channel VDAC". Nature. 399 (6735): 483–7. doi:10.1038/20959. PMID 10365962.
- ↑ Schwarzer C, Barnikol-Watanabe S, Thinnes FP, Hilschmann N (September 2002). "Voltage-dependent anion-selective channel (VDAC) interacts with the dynein light chain Tctex1 and the heat-shock protein PBP74". The International Journal of Biochemistry & Cell Biology. 34 (9): 1059–70. doi:10.1016/s1357-2725(02)00026-2. PMID 12009301.
- ↑ Kusano H, Shimizu S, Koya RC, Fujita H, Kamada S, Kuzumaki N, Tsujimoto Y (October 2000). "Human gelsolin prevents apoptosis by inhibiting apoptotic mitochondrial changes via closing VDAC". Oncogene. 19 (42): 4807–14. doi:10.1038/sj.onc.1203868. PMID 11039896.
- ↑ Baines CP, Song CX, Zheng YT, Wang GW, Zhang J, Wang OL, Guo Y, Bolli R, Cardwell EM, Ping P (May 2003). "Protein kinase Cepsilon interacts with and inhibits the permeability transition pore in cardiac mitochondria". Circulation Research. 92 (8): 873–80. doi:10.1161/01.RES.0000069215.36389.8D. PMC 3691672. PMID 12663490.
- ↑ Sun Y, Vashisht AA, Tchieu J, Wohlschlegel JA, Dreier L (November 2012). "Voltage-dependent anion channels (VDACs) recruit Parkin to defective mitochondria to promote mitochondrial autophagy". The Journal of Biological Chemistry. 287 (48): 40652–60. doi:10.1074/jbc.M112.419721. PMC 3504778. PMID 23060438.
- ↑ Noskov SY, Rostovtseva TK, Bezrukov SM (December 2013). "ATP transport through VDAC and the VDAC-tubulin complex probed by equilibrium and nonequilibrium MD simulations". Biochemistry. 52 (51): 9246–56. doi:10.1021/bi4011495. PMID 24245503.
Further reading
- Verrier F, Mignotte B, Jan G, Brenner C (December 2003). "Study of PTPC composition during apoptosis for identification of viral protein target". Annals of the New York Academy of Sciences. 1010 (1): 126–42. doi:10.1196/annals.1299.022. PMID 15033708.
- McEnery MW, Snowman AM, Trifiletti RR, Snyder SH (April 1992). "Isolation of the mitochondrial benzodiazepine receptor: association with the voltage-dependent anion channel and the adenine nucleotide carrier". Proceedings of the National Academy of Sciences of the United States of America. 89 (8): 3170–4. doi:10.1073/pnas.89.8.3170. PMC 48827. PMID 1373486.
- Dawson SJ, White LA (May 1992). "Treatment of Haemophilus aphrophilus endocarditis with ciprofloxacin". The Journal of Infection. 24 (3): 317–20. doi:10.1016/S0163-4453(05)80037-4. PMID 1602151.
- Jürgens L, Ilsemann P, Kratzin HD, Hesse D, Eckart K, Thinnes FP, Hilschmann N (July 1991). "Studies on human porin. IV. The primary structures of "Porin 31HM" purified from human skeletal muscle membranes and of "Porin 31HL" derived from human B lymphocyte membranes are identical". Biological Chemistry Hoppe-Seyler. 372 (7): 455–63. doi:10.1515/bchm3.1991.372.2.455. PMID 1657034.
- Kayser H, Kratzin HD, Thinnes FP, Götz H, Schmidt WE, Eckart K, Hilschmann N (December 1989). "[Identification of human porins. II. Characterization and primary structure of a 31-lDa porin from human B lymphocytes (Porin 31HL)]". Biological Chemistry Hoppe-Seyler. 370 (12): 1265–78. PMID 2559745.
- Brdiczka D, Kaldis P, Wallimann T (November 1994). "In vitro complex formation between the octamer of mitochondrial creatine kinase and porin". The Journal of Biological Chemistry. 269 (44): 27640–4. PMID 7525559.
- Yu WH, Wolfgang W, Forte M (June 1995). "Subcellular localization of human voltage-dependent anion channel isoforms". The Journal of Biological Chemistry. 270 (23): 13998–4006. doi:10.1074/jbc.270.23.13998. PMID 7539795.
- Thomas L, Blachly-Dyson E, Colombini M, Forte M (June 1993). "Mapping of residues forming the voltage sensor of the voltage-dependent anion-selective channel". Proceedings of the National Academy of Sciences of the United States of America. 90 (12): 5446–9. doi:10.1073/pnas.90.12.5446. PMC 46737. PMID 7685903.
- Blachly-Dyson E, Zambronicz EB, Yu WH, Adams V, McCabe ER, Adelman J, Colombini M, Forte M (January 1993). "Cloning and functional expression in yeast of two human isoforms of the outer mitochondrial membrane channel, the voltage-dependent anion channel". The Journal of Biological Chemistry. 268 (3): 1835–41. PMID 8420959.
- Schleiff E, Shore GC, Goping IS (March 1997). "Human mitochondrial import receptor, Tom20p. Use of glutathione to reveal specific interactions between Tom20-glutathione S-transferase and mitochondrial precursor proteins". FEBS Letters. 404 (2–3): 314–8. doi:10.1016/S0014-5793(97)00145-2. PMID 9119086.
- Reymann S, Haase W, Krick W, Burckhardt G, Thinnes FP (August 1998). "Endosomes: another extra-mitochondrial location of type-1 porin/voltage-dependent anion-selective channels". Pflügers Archiv. 436 (3): 478–80. doi:10.1007/s004240050659. PMID 9644232.
- Narita M, Shimizu S, Ito T, Chittenden T, Lutz RJ, Matsuda H, Tsujimoto Y (December 1998). "Bax interacts with the permeability transition pore to induce permeability transition and cytochrome c release in isolated mitochondria". Proceedings of the National Academy of Sciences of the United States of America. 95 (25): 14681–6. doi:10.1073/pnas.95.25.14681. PMC 24509. PMID 9843949.
- Crompton M, Virji S, Ward JM (December 1998). "Cyclophilin-D binds strongly to complexes of the voltage-dependent anion channel and the adenine nucleotide translocase to form the permeability transition pore". European Journal of Biochemistry. 258 (2): 729–35. doi:10.1046/j.1432-1327.1998.2580729.x. PMID 9874241.
- Messina A, Oliva M, Rosato C, Huizing M, Ruitenbeek W, van den Heuvel LP, Forte M, Rocchi M, De Pinto V (February 1999). "Mapping of the human Voltage-Dependent Anion Channel isoforms 1 and 2 reconsidered". Biochemical and Biophysical Research Communications. 255 (3): 707–10. doi:10.1006/bbrc.1998.0136. PMID 10049775.
- Shimizu S, Narita M, Tsujimoto Y (June 1999). "Bcl-2 family proteins regulate the release of apoptogenic cytochrome c by the mitochondrial channel VDAC". Nature. 399 (6735): 483–7. doi:10.1038/20959. PMID 10365962.
- Decker WK, Bowles KR, Schatte EC, Towbin JA, Craigen WJ (October 1999). "Revised fine mapping of the human voltage-dependent anion channel loci by radiation hybrid analysis". Mammalian Genome. 10 (10): 1041–2. doi:10.1007/s003359901158. PMID 10501981.
- Jacotot E, Ravagnan L, Loeffler M, Ferri KF, Vieira HL, Zamzami N, Costantini P, Druillennec S, Hoebeke J, Briand JP, Irinopoulou T, Daugas E, Susin SA, Cointe D, Xie ZH, Reed JC, Roques BP, Kroemer G (January 2000). "The HIV-1 viral protein R induces apoptosis via a direct effect on the mitochondrial permeability transition pore". The Journal of Experimental Medicine. 191 (1): 33–46. doi:10.1084/jem.191.1.33. PMC 2195797. PMID 10620603.
- Shimizu S, Tsujimoto Y (January 2000). "Proapoptotic BH3-only Bcl-2 family members induce cytochrome c release, but not mitochondrial membrane potential loss, and do not directly modulate voltage-dependent anion channel activity". Proceedings of the National Academy of Sciences of the United States of America. 97 (2): 577–82. doi:10.1073/pnas.97.2.577. PMC 15372. PMID 10639121.
- Stadtmüller U, Eben-Brunnen J, Schmid A, Hesse D, Klebert S, Kratzin HD, Hesse J, Zimmermann B, Reymann S, Thinnes FP, Benz R, Götz H, Hilschmann N (December 1999). "Mitochondria-derived and extra-mitochondrial human type-1 porin are identical as revealed by amino acid sequencing and electrophysiological characterisation". Biological Chemistry. 380 (12): 1461–6. doi:10.1515/BC.1999.189. PMID 10661876.
External links
- VDAC1+protein,+human at the US National Library of Medicine Medical Subject Headings (MeSH)