GC-gehalte
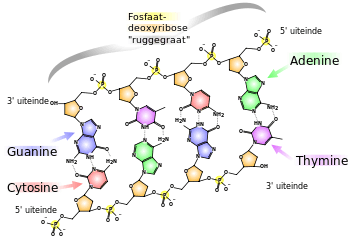
Het GC-gehalte is een kenmerk van DNA-moleculen en geeft het aandeel van de basen guanine (G) en cytosine (C) in het totaal van de basen (guanine, cytosine, adenine (A) en thymine (T)) in procenten weer.

GC-gehalte [%] = :
Een GC-gehalte van bijvoorbeeld 64 % betekent een AT-gehalte van 36 %.

Het GC-gehalte kan slaan op een speciaal fragment van het DNA of op het hele genoom. Wanneer het slaat op een fragment kan het het CG-gehalte zijn van een deel van een gen (domein), en enkel gen, groep van genen of zelfs van een niet-coderend gedeelte.
Guanine en cytosine zijn met elkaar verbonden door drie waterstofbruggen, terwijl adenine en thymine met twee bruggen zijn verbonden (G≡C versus A=T). DNA met een hoger GC-gehalte zou daardoor stabieler zijn dan dat met een laag GC-gehalte. De stabilisatie wordt echter hoofdzakelijk verkregen door de interacties van de stapelingen. De interacties van de stapelingen zijn energetisch het gunstigst voor gestapelde GC-GC en minder gunstig voor gestapelde AT-AT. Tussen de aromatische ringen van de heterocyclische basen bestaat een dipool-dipoolinteractie, die energetisch gunstig is. Dit effect verklaart voor een belangrijk gedeelte de grotere thermostabiliteit (hittebestendigheid) van DNA met een hoger GC-gehalte.[1] De vorming van het eerste basenpaar is op basis van de kleine energetisch winst en het verlies aan entropie nogal ongunstig, maar de verlenging van de helix met de volgende basenparen is door de stapeling wel energetisch gunstig.
Ondanks de grotere thermostabiliteit van het genetische materiaal, blijkt dat cellen met DNA met een hoog GC-gehalte autolyse vertonen, waardoor de levensduur van de cel juist verkort wordt.[2]
Taxonomie
Het GC-gehalte in het genoom wordt in de taxonomie gebruikt als een kenmerk voor de indeling van organismen, speciaal bij bacteriën. De gehalten bij de bacteriën lopen uiteen van ongeveer 20 tot bijna 80%. Bacteriën met een hoog GC-gehalte behoren vooral tot de straalzwammen (Actinobacteria), maar ook de Deltaproteobacteria zoals de Myxobacteria hebben een hoog GC-gehalte. Thermofiele organismen hebben ook een hoog GC-gehalte, wat terug te voeren is op de grotere stabiliteit van de G-C-basenparen.
Het GC-gehalte bij enkele modelorganismen:
| Soort | Phylogenetische groep | GC-gehalte |
|---|---|---|
| Streptomyces coelicolor Myxococcus xanthus Halobacterium sp. Saccharomyces cerevisiae (bakkersgist) Arabidopsis thaliana (zandraket) Methanosphaera stadtmanae Plasmodium falciparum (Malaria veroorzaker) |
straalzwammen (Actinobacteria) Deltaproteobacteria Archaea Ascomyceet (schimmel) Embryophyta Archaea Protozoa |
72 % 68 % 67 % 38 % 36 % 27 % ≈20 % |
Ter vergelijking: het GC-gehalte van het menselijk genoom bedraagt 41 % (zie ook CpG-eiland).
GC-gehalte van een DNA-gedeelte
Het aandeel basenparen GC en AT varieert ook binnen het genoom.
AT-rijke, dus GC-arme, gebieden in een genoom zitten vaak op die plaatsen waar de dubbele helix gemakkelijk uit elkaar moet kunnen gaan, bijvoorbeeld op die plaatsen waar de replicatie van het DNA begint. Ook bij mensen bestaan chromosomen met gebieden met een GC-gehalte dat duidelijk van 50 % afwijkt. Deze gebieden zijn meestal betrokken bij het instandhouden van de ruimtelijke structuur van de chromosomen.
Daarnaast is het GC-gehalte in DNA-gedeelten, die voor een gen coderen vaak hoger dan de gebieden met bijvoorbeeld introns of regulerende sequenties. Deze eigenschap wordt gebruikt voor het zoeken naar genen bij het sequencen van een genoom. Genoomsequencen bestaan uit miljoenen basen waartussen de open leesramen gevonden moeten worden. Het zoeken naar deze open leesramen wordt gedaan met behulp van computerprogramma's, zoals GLIMMER (Gene Locator and Interpolated Markov ModelER) bij bacteriën, archaea en virussen.[3][4]
Als er een functioneel gen (codogen) gevonden wordt waarvan het GC-gehalte duidelijk afwijkt van de overige genen, kan dat de oorzaak zijn van een nog niet zo lang geleden horizontale genoverdracht of van een gen dat afkomstig is van een retrovirus.
Bepaling van het GC-gehalte
Het GC-gehalte van DNA kan, indien de sequentie niet bekend is, experimenteel met verschillende methoden vastgesteld worden. De eenvoudigste methode is het meten van de denaturatietemperatuur (Tm-waarde) van de DNA-duppelstreng met behulp van een lichtmeter. Het DNA absorbeert ultraviolet licht van 260 nm. De dubbelstreng denatureert bij het verhitten in twee losse strengen, waardoor de lichtabsorptie met ongeveer 40% toeneemt. Dit effect wordt hyperchromiciteit genoemd.
De Tm-waarde wordt gedefinieerd als de temperatuur waarbij 50% van de dubbelstreng gedenatureerd is. De Tm-waarde van den DNA-dubbelstreng is direct afhankelijk van het GC-gehalte. Hoe meer GC-bindingen een DNA-molecuul heeft des te hoger is de Tm-waarde. Met de fotometrisch vastgestelde denaturatietemperatuur kan het GC-gehalte met de empirische formule (Tm [C°] – 69,4 C°) × 2,44 berekend worden.[5] De Tm-waarde is ook afhankelijk van de ionsterkte en het DNA-oplosmiddel. Daarom moet de denaturatietemperatuur vastgesteld worden in een gestandaardiseerde fysiologische zoutoplossing.
Veel nauwkeuriger is het vaststellen van het GC-gehalte met behulp van gaschromatografie.
Bronnen, noten en/of referenties
|