Replicatie (DNA)
Replicatie is het proces waarin DNA verdubbeld wordt. Het vindt plaats in alle levensvormen en is de basis voor de instandhouding van het erfelijk materiaal. Het DNA repliceert zich in de S-fase van de celcyclus. Pas wanneer een cel al zijn genetische informatie heeft gekopieerd kunnen bij de celdeling twee dochtercellen ontstaan, die dan beiden beschikken over een identiek genoom.

DNA replicatie is een complex proces waarbij veel verschillende eiwitten zijn betrokken.[1][2] Een hoofdrol speelt DNA-polymerase, die voor de polymerisatie van nucleotiden zorgt, zodat een nieuwe DNA-streng wordt gevormd.[3] De nucleotiden van de nieuwe streng worden verbonden met zogenaamde fosfodiësterbindingen (covalente bindingen tussen de OH-groep en de fosfaatgroep van de nucleotiden). In prokaryoten zijn drie DNA-polymerasen aangetoond: polymerase I, II en III. De replicatie van prokaryotisch DNA is goed onderzocht en voor een groot deel opgehelderd. Het kopiëren van DNA gebeurt met een precisie die aan het wonderbaarlijke grenst: DNA-polymerase heeft een intrinsieke foutmarge van minder dan één fout per tien miljoen toegevoegde nucleotiden.[4]
In het kernplasma bevinden zich onder andere de vrije nucleotiden dATP, dTTP, dGTP en dCTP. Dit zijn de bouwstenen die gebruikt worden bij DNA-synthese. Ze bestaan ieder uit desoxyribose (d), een base (A, T, G of C) en drie fosfaatgroepen (TP). Door twee fosfaatgroepen af te splitsen, komt chemische energie vrij. DNA-polymerase kan alleen aanhaken op een hydroxylgroep aan een 3’-eind. Om replicatie te beginnen, wordt een kort RNA-fragment, een zogenaamde primer, gekoppeld aan de template DNA-streng. Deze primer wordt gevormd door het enzym primase (een speciale RNA-polymerase).
Startpunt
DNA-replicatie begint bij een replicatiestartpunt of origin of replication. Deze startpunten zijn AT-rijke sequenties[kleine-letter 1] van ongeveer 250 basenparen lang. Adenine en thymine zijn verbonden met slechts twee waterstofbruggen. Meerdere van dit soort basenkoppelingen naast elkaar kunnen de dubbele streng lokaal verzwakken, waardoor het DNA hier gemakkelijker uit elkaar gaat. Bij eukaryoten bevat een DNA-molecuul veel replicatiestartpunten. Een DNA-molecuul van een prokaryoot heeft er maar één. Zodra het startpunt is gevonden door specifieke initiator-eiwitten, wordt een pre-replicatiecomplex gevormd, dat het dubbelstrengs DNA uit elkaar haalt. Vanuit de origin of replication verloopt replicatie in twee richtingen. Hierdoor ontstaat een replicatiebel die in breedte groeit. De replicatiebel kan gezien worden als een combinatie van twee replicatievorken.
Voordat een DNA-streng gerepliceerd kan worden, moet die zich eerst in een niet-gecondenseerde staat bevinden. DNA kan namelijk zeer dicht opgevouwen zijn rond histonen. Terwijl het DNA opgevouwen net te zien is onder de lichtmicroscoop, heeft het menselijke DNA, uit een cel, geheel ontvouwen een totale lengte van zo'n 2 meter, en slechts enkele nanometers in dikte.
Verloop
Initiatie
In twee richtingen worden de waterstofbruggen tussen de basenparen verbroken door het enzym helicase. De helixstructuur verdwijnt en de twee strengen gaan uit elkaar. Het maakt daarbij gebruik van de hydrolyse van ATP voor de benodigde energie. Het enzym topoïsomerase begeleidt het uit elkaar gaan van de ketens. Alle veranderingen van structuur leiden tot spanningsveranderingen en rotatiekrachten in de keten. Het topoïsomerase voorkomt dat dergelijke spanningen de keten beschadigen door de fosfodiësterbindingen voortdurend te verbreken en te vernieuwen. Op de plaats waar basenparing is verbroken, binden speciale eiwitten (single-strand binding proteins) aan de strengen. Deze eiwitten voorkomen dat de vrijgekomen basen opnieuw waterstofbruggen gaan vormen met de oude streng.
Elongatie
Het enzym DNA-polymerase III schuift vervolgens langs de enkelvoudige (enkelstrengse) ketens en bindt dATP, dTTP, dGTP en dCTP uit het kernplasma aan de vrijgekomen stikstofbasen, beginnende bij het 3’-eind van de primer. Hiervoor wordt energie gebruikt die vrijkomt door het afpitsen van twee fosfaatgroepen. Er ontstaan twee dubbelstrengs DNA-moleculen die ieder uit een oude en een nieuwe keten bestaan. Het DNA-polymerase leest de oorspronkelijke DNA-streng af van het 3’-eind naar het 5’-eind, waardoor de nieuwe DNA-streng wordt gesynthetiseerd van het 5’-eind naar het 3’-eind. Een volgende nucleotide kan alleen binden aan het 3’-eind van een nucleotide dat al is ingebouwd. Langs beide nucleotideketens bewegen de DNA-polymerase-enzymen zich in tegengestelde richting om een nieuwe keten te synthetiseren.
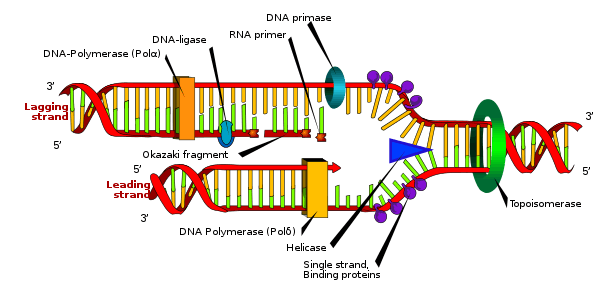
In de ene richting kan DNA-polymerase vanaf het replicatiestartpunt het uit elkaar gaan van de ketens volgen om de leading strand te vormen. In de andere richting kan DNA-polymerase steeds maar kleine stukjes synthetiseren, doordat dit achterwaarts moet gebeuren: nucleotiden koppelen alleen aan het 3’-eind van de RNA-primer. Primase zet om zoveel basen een primer af, waartussen nucleotiden worden ingebouwd. Deze stukjes zijn ongeveer 100 tot 200 nucleotiden lang en worden Okazaki-fragmenten genoemd. Het enzym DNA-ligase koppelt de ze aan elkaar waardoor de lagging strand wordt gevormd.
Terminatie
DNA-polymerase I vervangt uiteindelijk de primer door DNA. De ribonucleotiden van de primer worden daarbij verwisseld voor desoxyribonucleotiden. Het nieuw gevormde nucleïnezuur wordt geïnspecteerd door een aantal controle-eiwitten die alle lassen in de gevormde keten nog eens nalopen op kwaliteit. Zij voeren eventuele correcties uit.
Op plaatsen waar een nieuwe complementaire nucleotide-keten is ontstaan, neemt het DNA weer een helixstructuur aan. Na spiralisatie van het DNA bestaat het chromosoom uit twee chromatiden. Op de plaats van de centromeer worden beide chromatiden nog door waterstofbruggen bij elkaar gehouden. Tijdens de mitose kunnen ze uit elkaar gaan.
Telomeren
Iedere keer dat het DNA op de gefragmenteerde wijze wordt gedupliceerd zoals bij de lagging strand, wordt deze streng iets korter. Op een gegeven moment is er namelijk geen ruimte meer om nog een laatste primer te plaatsen die het uiteinde voorziet van polymerisatie. Deze verkorting treedt op bij de telomeer. Gelukkig bevatten telomeren geen coderende regio’s, maar het aantal delingen is toch beperkt. Er komt een moment dat verdergaande verkorting van de uiteinden van het DNA-molecuul leidt tot het verlies van kritische informatie. In voortplantingscellen en stamcellen komt een enzym voor dat voorkomt dat het telomeer korter wordt: telomerase. Telomerase voegt na iedere DNA replicatie de specifiek telomeer-basenvolgorde toe. Het telomeer wordt zo nooit korter en de cel kan blijven delen, zonder dat kritische informatie verloren gaat.
Zie ook
| Noten
Referenties
|
| DNA-replicatie (eukaryoten) | ||||||||
|---|---|---|---|---|---|---|---|---|
|