Genetic history of North Africa

The genetic history of North Africa has been heavily influenced by geography. The Sahara desert to the south and the Mediterranean Sea to the North were important barriers to gene flow in prehistoric times. However, Northeast Africa and the Levant form a single land mass at the Suez. At the Straits of Gibraltar, North Africa and Europe are separated by only 15 km (9 mi).
Although North Africa has experienced gene flows from the surrounding regions, it has also experienced long periods of genetic isolation, allowing a distinctive genetic "Berber marker" to evolve in the native Berber people. Today, this genetic "Berber marker" is consistently found in the regions and populations that still predominantly speaking the Berber language, as well as in the Canary Islands which was inhabited by native Berbers and by their descendents to this day. A recent genetic study showed that North Africans are genetically similar to Paleolithic North Africans.[1]
Current scientific debate is concerned with determining the relative contributions of different periods of gene flow to the current gene pool of North Africans. Anatomically modern humans are known to have been present in North Africa during the Middle Paleolithic (300,000 years ago), as attested by the by Jebel Irhoud 1.[2] With no apparent continuity, 25,000 years ago, the Aterian was succeeded by the Iberomaurusian industry, whose lithic assemblages bore relations with the Cro-Magnon cultures. The Iberomaurusian industry was succeeded by the Capsian industry in the eastern part of North Africa.
In the 7th century A.D., part of the Berber countries was invaded by Muslim Umayyad Arabs. Under the relatively brief Arab-Umayyad occupation and the later arrival of some bedouin Arabs and Syriacs from the Near East in Asia and the arrival of some Jews and Muslims fleeing the Spanish Catholic Reconquista, a partial population mix or fusion might have taken place and might have resulted in some genetic diversity among some North Africans.[3] However, this partial fusion of Berbers and foreigners is mostly limited in terms of geographical distribution to the main Berber urban areas and some coastal plains of North Africa because migrants and refugees tend to gravitate towards major cities since ancient times and they tend to avoid the heartland. The Berber ethnic and genetic nature of North Africa (west of Egypt) is still dominant, either prominently (as in language or ethnic identity) or subtly (as in culture and genetic heritage).
Y-chromosome
Haplogroup E is the most common paternal haplogroup among Berbers. It represents up to 100 percent of Y-chromosomes among some Berber populations. Haplogroup E is thought to have emerged in prehistoric North Africa or East Africa,[4] and would have later dispersed into West Asia. The major subclades of haplogroup E found amongst Berbers belong to E-Z827, which is believed to have emerged in North Africa. Common subclades include E1b1b1a, E1b1b1b and E1b1b1*. E1b1b1b is distributed along a west-to-east cline with frequencies that can reach as high as 100 percent in Northwest Africa. E1b1b1a has been observed at low to moderate frequencies among Berber populations with significantly higher frequencies observed in Northeast Africa relative to Northwest Africa.[5][6][7]
West Eurasian haplogroups, such as Haplogroup J and Haplogroup R1, have also been observed at moderate frequencies. A thorough study by Arredi et al. (2004), which analyzed populations from Algeria, concludes that the North African pattern of Y-chromosomal variation (including both J1 and E1b1b main haplogroups) is largely of Neolithic origin, which suggests that the Neolithic transition in this part of the world was accompanied by demic diffusion of Berber–speaking pastoralists from the Middle East[5][8] However, Loosdrecht et al. 2018 demonstrated that E1b1b is most likely indigenous to North Africa and migrated from North Africa to the Near East during the Paleolithic.[1]
E1b1b1b (E-M81); formerly E3b1b, E3b2

E1b1b1b (E-M81) is the most common Y chromosome haplogroup in North Africa, dominated by its sub-clade E-M183. It is thought to have originated in North Africa 5,600 years ago. The parent clade, E1b1b, originated in East Africa.[4][9] Colloquially referred to as the Berber marker or Maghrebi marker for its prevalence among Mozabite, Middle Atlas, and other Berber-speaking groups, E-M81 is also quite common among North African groups. It reaches frequencies of up to 90 percent in some parts of the Maghreb. This includes the Saharawish for whose men Bosch et al. (2001) reports that approximately 76 percent are M81+.
This haplogroup is also found at high levels in Canary islands and parts of the Iberian Peninsula (Spain and Portugal) as well as much lower levels in Italy and France. In Iberia, it is generally more common than E1b1b1a (E-M78),[10] unlike the rest of Europe, and as a result E-M81 is found throughout Latin America[11] and among Hispanic men in USA.[12] As an exceptional case in Europe, this sub-clade of E1b1b1 has also been observed at 40 percent the Pasiegos from Cantabria.[4]
Another theory by Gonçalves suggests that, since the percentage of E in Iberia is high in areas with little to no Moorish/Berber settlement, such as Galicia, Castile and Cantabria, the lineage may have been brought to Iberia by Neolithic or Mesolithic migrations. The cause for such migrations may have been the drying out of the Sahara and the movement of populations from Northwest Africa across the Strait of Gibraltar northward to more temperate Iberia. This theory suggests a pre-Islamic prehistoric migration from North Africa into Iberia that has left behind a genetic trace in the modern Iberian population. More research is required in determining how close the subclades of E carried by Iberian men are related to Maghrebis, and whether they are exactly the same subclade. This would help to determine whether E-M81's presence in Iberia is prehistoric or from more recent periods, such the Islamic civilization of Al-Andalus.
In smaller numbers, E-M81 men can be found in Sudan, Lebanon, Turkey, and among Sephardic Jews.
There are two recognized sub-clades, although one is much more common than the other.
- Sub clades of E1b1b1b (E-M81):
- E1b1b1b1 (E-M107). Underhill et al. (2000) found one example in Mali.
- E1b1b1b2 (E-M183). Individuals with the defining marker for this clade, M81, also test positive, in tests so far, for M183. As of October 23, 2008, the SNP M165 is currently considered to define a subclade, "E1b1b1b2a".[13]
Mitochondrial DNA


Individuals receive mtDNA only from their mothers. According to Macaulay et al. 1999, "one-third (33%) of Mozabite Berber mtDNAs have a Near Eastern ancestry, probably having arrived in North Africa less than 50,000 years ago, and one-eighth (12.5%) have an origin in sub-Saharan Africa. Europe appears to be the source of many of the remaining sequences, with the rest having arisen either in Europe or in the Near East".[14] Maca-Meyer et al. 2003 analyze the "autochthonous North African lineage U6" in mtDNA, and conclude that:
- The most probable origin of the proto-U6 lineage was the Near East. Around 30,000 years ago it spread to North Africa where it represents a signature of regional continuity. Subgroup U6a reflects the first North African expansion from the Maghreb returning to the east in Paleolithic times. Derivative clade U6a1 signals a posterior movement from Northeast Africa back to the Maghreb and the Near East. This migration coincides with a possible
Afroasiatic linguistic expansion.
A genetic study by Fadhlaoui-Zid et al. 2004[15] argues concerning certain exclusively North African haplotypes that "expansion of this group of lineages took place around 10,500 years ago in North Africa, and spread to neighbouring population", and apparently that a specific Northwestern African haplotype, U6, probably originated in the Near East 30,000 years ago accounts for 28 percent in Mozabites, 18 percent in Kabyles, but has not been preserved in the southern Moroccan Berbers and accounts for 6-8 percent. Rando et al. 1998 (as cited by ) "detected female-mediated gene flow from sub-Saharan Africa to NW Africa" amounting to as much as 21.5 percent of the mtDNA sequences in a sample of NW African populations; the amount varied from 82 percent in Tuaregs to less than 3 percent in Riffians in north of Morocco. This north-south gradient in the sub-Saharan contribution to the gene pool is supported by Esteban et al.[16]
Nevertheless, individual Berber communities display a considerably high mtDNA heterogeneity among them. The Berbers of Jerba Island, located in South Eastern Tunisia, display an 87 percent West Eurasian contribution with no U6 haplotypes,[17] while the Kesra of Tunisia, for example, display a much higher proportion of typical sub-Saharan mtDNA haplotypes (49 percent),[18] as compared to the Zriba (8 percent). According to the article, "The North African patchy mtDNA landscape has no parallel in other regions of the world and increasing the number of sampled populations has not been accompanied by any substantial increase in our understanding of its phylogeography. Available data up to now rely on sampling small, scattered populations, although they are carefully characterized in terms of their ethnic, linguistic, and historical backgrounds. It is therefore doubtful that this picture truly represents the complex historical demography of the region rather than being just the result of the type of samplings performed so far."
Haplotype V, have its highest concentration among the Saami people of northern Scandinavia (approximately 59%). It has been found at approximately 10% among the Mari people of the Volga-Ural region, leading to the suggestion that this region might be the source of the V among the Saami.
Haplogroup V is also found at higher than average levels in Cantabrian people (15%) of northern Iberia, and somewhat lower in nearby Basque people (10.4%). It also is found in particularly high concentrations (16.3%) among the Berbers of Matmata, Tunisia. The frequency of haplotype V is to be observed throughout the Mediterranean region, ranging from frequencies of close to 30 percent in southern Portugal to around 10 percent in southern France. Similarly, the highest frequency in Italy is to be found in the southern island of Sicily (28 percent).[19][20]
Additionally, recent studies have discovered a close mitochondrial link between Berbers and the Saami of Scandinavia, which confirms that Southwestern Europe and North Africa was the source of late-glacial expansions of hunter-gatherers that repopulated Northern Europe after the Last Glacial Maximum and reveals a direct maternal link between those European hunter-gatherer populations and the Berbers.[18][21] With regard to Mozabite Berbers, one-third (33%) of Mozabite Berber mtDNAs have a Near Eastern ancestry, probably having arrived in North Africa ∼50,000 years ago, and one-eighth (12.5%) have an origin in sub-Saharan Africa. Europe appears to be the source of many of the remaining sequences, with the rest (54.5%) having arisen either in Europe or in the Near East."[22]
According to the most recent and thorough study on Berber mtDNA from Coudray et al. 2008, which analysed 614 individuals from 10 different regions (Morocco (Asni, Bouhria, Figuig, Souss), Algeria (Mozabites), Tunisia (Chenini-Douiret, Sened, Matmata, Jerba) and Egypt (Siwa)),[23] the results may be summarized as follows:
- Total West Eurasian lineages (H, HV, R0, J, M, T, U, K, N1, N2, X) : 80 percent
- Total African lineages (L0, L1, L2, L3, L4, L5) : 20 percent
The Berber mitochondrial pool is characterized by an overall high frequency of Western Eurasian haplogroups, a somehow lower frequency of sub-Saharan L lineages, and a significant (but differential) presence of North African haplogroups U6 and M1.
Some papers suggest that the distribution of the main L haplogroups in North Africa was mainly due to trans-Saharan slave trade.[24] However, in September 2010, a thorough study of Berber mtDNA by Frigi et al. concluded that most of L haplogroups were much older and introduced by an ancient African gene flow around 20,000 years ago.[25]
Autosomal DNA
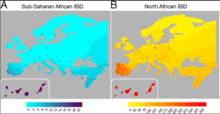
On 13 January 2012, an exhaustive genetic study of North Africa's human populations was published in PLoS Genetics and was undertaken jointly by researchers in the Evolutionary Biology Institute (CSIC-UPF) and Stanford University, among other institutions.[26]
The study reveals that the genetic composition of North Africa's human populations is extremely complex, and the result of a local component dating back thirteen thousand years and the varied genetic influence of neighbouring populations on North African groups during successive migrations. According to David Comas, coordinator of the study and researcher at the Institute for Evolutionary Biology (CSIC-UPF), "some of the questions we wanted to answer were whether today's inhabitants are direct descendants of the populations with the oldest archaeological remains in the region, dating back fifty thousand years, or whether they are descendants of the Neolithic populations in the Middle East, which introduced agriculture to the region around eight thousand years ago. We also wondered if there had been any genetic exchange between the North African populations and the neighbouring regions and if so, when these took place".
To answer these questions, the researchers analyzed around 800,000 genetic markers, distributed throughout the entire genome in 125 North African individuals belonging to seven representative populations in the whole region, and the information obtained was compared with the information from the neighbouring populations.
The results of this study show that there is a native genetic component that defines North Africans. In-depth study of these markers shows that the people inhabiting North Africa today are not descendants of the earliest occupants of this region fifty thousand years ago, but shows that the ancestors of today's North Africans were a group of populations that already lived in the region around thirteen thousand years ago. Furthermore, this local North African genetic component is very different from the one found in the populations in the south of the Sahara, which shows that the ancestors of today's North Africans were members of a subgroup of humanity who left Africa to conquer the rest of the world and who subsequently returned to the north of the continent to settle in the region.
As well as this local component, North African populations were also observed to share genetic markers with all the neighbouring regions, as a result of more recent migrations, although these appear in different proportions.
There is an influence from the Middle East, which becomes less marked as the distance from the Arabian Peninsula increases, similar proportions of European influence in all North African populations, and, in some populations, there are even individuals who present a large proportion of influence from the South of the Sahara in their genome.
A 2015 study by Dobon et al. identified an ancestral autosomal component of West Eurasian origin that is common to many modern Afro-Asiatic-speaking populations in Northeast Africa. Known as the Coptic component, it peaks among Egyptian Copts who settled in Sudan over the past two centuries. The Coptic component evolved out of a main North African and Middle Eastern ancestral component that is shared by other Egyptians and also found at high frequencies among other Afro-Asiatic populations in Northeast Africa (~70%). The scientists suggest that this points to a common origin for the general population of Egypt. They also associate the Coptic component with Ancient Egyptian ancestry, without the later Arabian influence that is present among other Egyptians.[27]
According to a paper published in 2017 :
Most of the genetic studies in North African populations agree with a limited or non-existent correlation between genetics and geography, and therefore, they show a high population heterogeneity in the region (Bosch et al., 2000; Fadhlaoui-Zid et al., 2004; Flores et al., 2011; González-Pérez et al., 2010). However, an east-west genetic pattern of variation has been observed (Arauna et al., 2017; Bosch et al., 1997; Harich et al., 2001; Henn et al., 2012), which has been explained by Middle Eastern gene flow towards North Africa according to genome-wide analyses (Arauna et al., 2017; Henn et al., 2012). The genetic heterogeneity is not only due to the wide extension of North Africa, since it is also found at microgeographical level when considering more restricted areas, such as Algeria (Bekada et al., 2015) or Tunisia (Fadhlaoui-Zid et al., 2011a; Kefi et al., 2015), for instance. Regardless of the genetic markers analysed, North African populations have been described as a mosaic of North African, Middle Eastern, European and sub-Saharan ancestries. The differential admixture with these four ancestry sources explains the current genetic structure in North Africa, characterised by diverse and heterogeneous populations. This differential admixture explains why nearby populations or even individuals inhabiting the same location might be genetically more distant than groups of people in geographically distant populations. For instance, North African individuals with similar amounts of sub-Saharan admixture present high genetic similarities regardless of their geographical origin. Neither geography, linguistics nor ethnic affiliation is correlated with a specific pattern of admixture proportions. A completely different scenario was shown when analysing North African Jews, who are genetically closer to other Jewish populations than to their geographic neighbours (Campbell et al., 2012). There are many possible scenarios that might explain the differential admixture in North Africa, socio-cultural structure and historical events being the main reasons. However, more complex socio-cultural scenarios, including socio-economic factors, a more precise ethnic or linguistic definition, particularities of the history of each local area and rural versus urban lifestyles, should be considered in order to find a correlation with the complex genetic pattern observed (Arauna et al., 2017).
— Arauna, Lara R and Comas, David (September 2017) Genetic Heterogeneity between Berbers and Arabs. In: eLS. John Wiley & Sons, Ltd: Chichester. DOI: 10.1002/9780470015902.a0027485
Genetic influence
Y-chromosome DNA
The general parent Y-chromosome Haplogroup E1b1b (formerly known as E3b), which might have originated in the Horn of Africa or the Near East[5] is by far the most common clade in North and Northeast Africa and found in select populations in Europe, particularly in the Mediterranean and South Eastern Europe. E1b1b reaches in Europe Greece and the Balkan region but, is not as high there as it is among African populations..[5]
{kind=link}
Outside of North and Northeast Africa, E1b1b's two most prevalent clades are E1b1b1a (E-M78, formerly E3b1a) and E1b1b1b (E-M81, formerly E3b1b).
The general parent Y-chromosome Haplogroup E1b1b (formerly known as E3b), which might have originated in North Africa, the Horn of Africa or the Near East[5] is by far the most common clade in North and Northeast Africa and found in select populations in Europe, particularly in the Mediterranean and South Eastern Europe. E1b1b reaches in Europe Greece and the Balkan region but, is not as high there as it is among African populations..[5]
A study from Semino (published 2004) showed that Y-chromosome haplotype E1b1b1b (E-M81), is specific to North African populations and almost absent in Europe except the Iberia (Spain and Portugal) and Sicily.[5] Another 2004 study showed that E1b1b1b is found present, albeit at low levels throughout Southern Europe (ranging from 1.5 percent in Northern Italians, 2.2 percent in Central Italians, 1.6 percent in Southern Spaniards, 3.5 percent in the French, 4 percent in the Northern Portuguese, 12.2 percent in the Southern Portuguese and 41.2 percent in the genetic isolate of the Pasiegos from Cantabria).[28]
The findings of this latter study contradict a more thorough analysis Y-chromosome analysis of the Iberian peninsula according to which haplogroup E1b1b1b surpasses frequencies of 10 percent in Southern Spain. The study points only to a very limited influence from Northern Africa and the Middle East in Iberia, both in historic and prehistoric times.[29] The absence of microsatellite variation suggests a very recent arrival from North Africa consistent with historical exchanges across the Mediterranean during the period of Islamic expansion, namely of Berber populations.[5] However, a study restricted to Portugal, concerning Y-chromosome lineages, revealed that "The mtDNA and Y-DNA data indicate that the presence of Berbers in that region dates clearly prior to the Moorish expansion in 711 AD, so it´s not recent there at all. ... Our data indicate that male Berbers, unlike sub-Saharan immigrants, constituted long-lasting and continuous community in the country".[30]
A wide-ranging study (published 2007) using 6,501 unrelated Y-chromosome samples from 81 populations found that: "Considering both these E-M78 sub-haplogroups (E-V12, E-V22, E-V65) and the E-M81 haplogroup, the contribution of northern African lineages to the entire male gene pool of Iberia (barring Pasiegos), continental Italy and Sicily can be estimated as 5.6 percent, 3.6 percent and 6.6 percent, respectively."[20] It has also been argued that the European distribution of E-M78 and its sub-clades is compatible with the Neolithic demic diffusion of agriculture, but also possibly partly from at least, the Mesolithic. For example, Battaglia et al. (2007) estimated that E-M78 (called E1b1b1a1 in that paper) has been in Europe longer than 10,000 years. In support of this theory, human remains excavated in a Spanish funeral cave dating from approximately 7,000 years ago were shown to be in this haplogroup.[31] More recently, two E-M78 have been found in the Neolitich Sopot and Lengyel cultures from the same period.[32]
High-resolution analysis of human Y-chromosome variation shows a sharp discontinuity and limited gene flow between northwestern Africa and the Iberian Peninsula (Bosch E, Calafell F, Comas D, Oefner PJ, Underhill PA, Bertranpetit J.) which seems supported by the most recent studies (including autosomal research).
A very recent study about Sicily by Gaetano et al. 2008 found that "The Hg E3b1b-M81, widely diffused in northwestern African populations, is estimated to contribute to the Sicilian gene pool at a rate of 6 percent." .[33]
According to the most recent and thorough study about Iberia by Adams et al. 2008 that analysed 1,140 unrelated Y-chromosome samples in Iberia, a limited contribution of northern African lineages to the entire male gene pool of Iberia was found : "mean North African admixture is just 10.6 percent, with wide geographical variation, ranging from zero in Gascony to 21.7 percent in Northwest Castile".[34][35] More recent extensive/complete studies, like «The Geography of Recent Genetic Ancestry across Europe», determined that Italians and Iberians, in fact, share very few common ancestors with other populations over at least, the last 2.500 years, unlike all the other European populations, present on the study, so North African contribution in both peninsulas is very likely limited (always a minority even in «hotspot» areas), many times constituted by ancient haplogroups, and in many cases, geographically not compatible with Moor invasion.
Mitochondrial DNA
Genetic studies on Iberian populations also show that North African mitochondrial DNA sequences (haplogroup U6) and sub-Saharan sequences (Haplogroup L), although present at only low levels, are still at higher levels than those generally observed elsewhere in Europe, though very likely, most of the L mtDNA that has been found in minor amounts in Iberia, is actually pre-neolithic in origin, as it was demonstrated by María Cerezo et al., (Reconstructing ancient mitochondrial DNA links between Africa and Europe) and U6 too, which also have a very old presence in Iberia, since Iberia has a great diversity in lineages from this haplogroup, it was already found in some local hunter-gatherer remains and its local geographic distribution is not compatible, in many cases, with Moor occupation area.[36][37][38] Haplogroup U6 have also been detected in Sicily and Southern Italy at much lower frequencies.[39] It happens also to be a characteristic genetic marker of the Saami populations of Northern Scandinavia.[21]
It is difficult to ascertain that U6's presence is the consequence of Islam's expansion into Europe during the Middle Ages, particularly because it is more frequent in the west of the Iberian Peninsula rather than in the east. In smaller numbers it is also attested in the British Isles, again in its northern and western borders. It may be a trace of a prehistoric Neolithic/Megalithic/Mesolithic or even Upper Paleolithic expansion along the Atlantic coasts from North Africa or Iberian Peninsula, perhaps in conjunction with seaborne trade, although an alternative, but less likely explanation, would attribute this distribution in Northern Britain to the Roman period. One subclade of U6 is particularly common among Canarian Spaniards as a result of native Guanche (proto-Berber) ancestry.
Genetic influences on Latin America
As a consequence of Spanish and Portuguese colonization of Latin America, E-M81 is also found throughout Latin America[40][41][42] and among Hispanic men in USA.[43]
Ancient DNA
In 2013, Nature announced the publication of the first genetic study utilizing next-generation sequencing to ascertain the ancestral lineage of an Ancient Egyptian individual. The research was led by Carsten Pusch of the University of Tübingen in Germany and Rabab Khairat, who released their findings in the Journal of Applied Genetics. DNA was extracted from the heads of five Egyptian mummies that were housed at the institution. All the specimens were dated to between 806 BC and 124 AD, a timeframe corresponding with the Late Dynastic and Ptolemaic periods. The researchers observed that one of the mummified individuals likely belonged to the mtDNA haplogroup I2, a maternal clade that is believed to have originated in Western Asia.[44]
In 2013, Iberomaurusian skeletons from the prehistoric sites of Taforalt and Afalou in the Maghreb were analyzed for ancient DNA. All of the specimens belonged to maternal clades associated with either North Africa or the northern and southern Mediterranean littoral, indicating gene flow between these areas since the Epipaleolithic.[45] The ancient Taforalt individuals carried the mtDNA haplogroups U6, H, JT and V, which points to population continuity in the region dating from the Iberomaurusian period.[46]
The E1b1b-M81 (~44%), R-M269 (~44%), and E-M132/E1a (~6%) paternal haplogroups have been found in ancient Guanche (Bimbapes) fossils excavated in Punta Azul, El Hierro, Canary Islands, which are dated to the 10th century. Maternally, the specimens all belong to the H1 clade. These locally born individuals carried the H1-16260 haplotype, which is exclusive to the Canary Islands and Algeria. Analysis of their autosomal STRs indicates that they were most closely related to Moroccan Berbers.[47]
In 2018, DNA analysis of Later Stone Age individuals from the site of Taforalt (Iberomaurusian, 15 000 BP) and Early Neolithic Moroccans from the site of Ifri N' Ammar (7 000 BP) revealed that they were related to the modern North Africans and carried Y-DNA E-M35, EM-215*, E-L19*, and E-M78*. These studies confirmed a long-term genetic continuity in the region showing that Mesolithic Moroccans are similar to Later Stone Age individuals from the same region and possess an endemic component retained in present-day Maghrebi populations (representing 20% to 70% of their total ancestry).[1][48]
See also
References
- 1 2 3 Loosdrecht, Marieke van de; Bouzouggar, Abdeljalil; Humphrey, Louise; Posth, Cosimo; Barton, Nick; Aximu-Petri, Ayinuer; Nickel, Birgit; Nagel, Sarah; Talbi, El Hassan (2018-03-15). "Pleistocene North African genomes link Near Eastern and sub-Saharan African human populations". Science: eaar8380. doi:10.1126/science.aar8380. ISSN 0036-8075. PMID 29545507.
- ↑ Callaway, Ewen (2017-06-07). "Oldest Homo sapiens fossil claim rewrites our species' history". Nature. doi:10.1038/nature.2017.22114. ISSN 1476-4687.
- ↑ Rando (1998), "Mitochondrial DNA analysis of Northwest African populations reveals genetic exchanges with European, Near-Eastern, and sub-Saharan populations" (PDF), Annals of Human Genetics, 62: 531–50, doi:10.1046/j.1469-1809.1998.6260531.x, PMID 10363131
- 1 2 3 Cruciani, F.; La Fratta, R.; Santolamazza, P.; Sellitto, D.; Pascone, R.; Moral, P.; Watson, E.; Guida, V.; Colomb, E. B. (2004). "Phylogeographic Analysis of Haplogroup E3b (E-M215) Y Chromosomes Reveals Multiple Migratory Events Within and Out of Africa". The American Journal of Human Genetics. 74 (5): 1014–1022. doi:10.1086/386294. PMC 1181964. PMID 15042509.
- 1 2 3 4 5 6 7 8 Semino, O.; Magri, C.; Benuzzi, G.; Lin, A. A.; Al-Zahery, N.; Battaglia, V.; MacCioni, L.; Triantaphyllidis, C.; Shen, P. (2004). "Origin, Diffusion, and Differentiation of Y-Chromosome Haplogroups E and J: Inferences on the Neolithization of Europe and Later Migratory Events in the Mediterranean Area". The American Journal of Human Genetics. 74 (5): 1023–34. doi:10.1086/386295. PMC 1181965. PMID 15069642.
- ↑ Kujanová, M.; Pereira, L. S.; Fernandes, V. N.; Pereira, J. B.; čErný, V. (2009). "Near Eastern Neolithic genetic input in a small oasis of the Egyptian Western Desert". American Journal of Physical Anthropology. 140 (2): 336–346. doi:10.1002/ajpa.21078. PMID 19425100.
- ↑ Fadhlaoui-Zid, Karima; Martinez-Cruz, Begoña; Khodjet-el-khil, Houssein; Mendizabal, Isabel; Benammar-Elgaaied, Amel; Comas, David (October 2011). "Genetic structure of Tunisian ethnic groups revealed by paternal lineages". American Journal of Physical Anthropology. 146 (2): 271–280. doi:10.1002/ajpa.21581. ISSN 1096-8644. PMID 21915847.
- ↑ although later papers have suggested that this date could have been as long as ten thousand years ago, with the transition from the Oranian to the Capsian culture in North Africa. SpringerLink - Journal Article
- ↑ Arredi et al. (2004)
- ↑ See for example Flores et al. (2004).
- ↑ See the remarks of genetic genealogist Robert Tarín for example. We can add 6.1 percent (8 out of 132) in Cuba, Mendizabal et al. (2008); 5.4 percent (6 out of 112) in Brazil (Rio de Janeiro), "The presence of chromosomes of North African origin (E3b1b-M81; Cruciani et al., 2004) can also be explained by a Portuguese-mediated influx, since this haplogroup reaches a frequency of 5.6 percent in Portugal (Beleza et al., 2006), quite similar to the frequency found in Rio de Janeiro (5.4 percent) among European contributors.", Silva et al. (2006)
- ↑ 2.4 percent (7 out of 295) among Hispanic men from California and Hawaii, Paracchini et al. (2003)
- ↑ Y-DNA Haplogroup E and its Subclades - 2008
- ↑ http://www.stats.gla.ac.uk/~vincent/papers/980656.web.pdf
- ↑ Fadhlaoui-Zid, K.; Plaza, S.; Calafell, F.; Ben Amor, M.; Comas, D.; Bennamar, A.; Gaaied, E. (2004). "Mitochondrial DNA Heterogeneity in Tunisian Berbers". Annals of Human Genetics. 68 (3): 222–33. doi:10.1046/j.1529-8817.2004.00096.x. PMID 15180702.
- ↑ Esteban, E.; González-Pérez, E.; Harich, N.; López-Alomar, A.; Via, M.; Luna, F.; Moral, P. (2004). "Genetic relationships among Berbers and South Spaniards based on CD4 microsatellite/Alu haplotypes". Annals of Human Biology. 31 (2): 202–212. doi:10.1080/03014460310001652275. PMID 15204363.
- ↑ Loueslati, B. Y.; Cherni, L.; Khodjet-Elkhil, H.; Ennafaa, H.; Pereira, L. S.; Amorim, A. N.; Ben Ayed, F.; Ben Ammar Elgaaied, A. (2006). "Islands Inside an Island: Reproductive Isolates on Jerba Island". American Journal of Human Biology. 18 (1): 149–153. doi:10.1002/ajhb.20473. PMID 16378336.
- 1 2 Cherni, L.; Loueslati, B. Y.; Pereira, L.; Ennafaa, H.; Amorim, A.; Gaaied, A. B. A. E. (2005). "Female Gene Pools of Berber and Arab Neighboring Communities in Central Tunisia: Microstructure of mtDNA Variation in North Africa". Human Biology. 77 (1): 61–70. doi:10.1353/hub.2005.0028. PMID 16114817.
- ↑ Gérard, N.; Berriche, S.; Aouizérate, A.; Diéterlen, F.; Lucotte, G. R. (2006). "North African Berber and Arab Influences in the Western Mediterranean Revealed by Y-Chromosome DNA Haplotypes". Human Biology. 78 (3): 307–316. doi:10.1353/hub.2006.0045. PMID 17216803.
- 1 2 Cruciani, F.; La Fratta, R.; Trombetta, B.; Santolamazza, P.; Sellitto, D.; Colomb, E. B.; Dugoujon, J. -M.; Crivellaro, F.; Benincasa, T. (2007). "Tracing Past Human Male Movements in Northern/Eastern Africa and Western Eurasia: New Clues from Y-Chromosomal Haplogroups E-M78 and J-M12". Molecular Biology and Evolution. 24 (6): 1300–1311. doi:10.1093/molbev/msm049. PMID 17351267.
- 1 2 Achilli, A.; Rengo, C.; Battaglia, V.; Pala, M.; Olivieri, A.; Fornarino, S.; Magri, C.; Scozzari, R.; Babudri, N. (2005). "Saami and Berbers—An Unexpected Mitochondrial DNA Link". The American Journal of Human Genetics. 76 (5): 883–886. doi:10.1086/430073. PMC 1199377. PMID 15791543.
- ↑ MacAulay, V.; Richards, M.; Hickey, E.; Vega, E.; Cruciani, F.; Guida, V.; Scozzari, R.; Bonné-Tamir, B.; Sykes, B.; Torroni, A. (1999). "The Emerging Tree of West Eurasian mtDNAs: A Synthesis of Control-Region Sequences and RFLPs". The American Journal of Human Genetics. 64 (1): 232–49. doi:10.1086/302204. PMC 1377722. PMID 9915963.
- ↑ Data from Achilli et al. 2005; Brakez et al. 2001; Cherni et al. 2005; Fadhlaoui-Zid et al. 2004; Krings et al.1999; Loueslati et al. 2006; Macaulay et al. 1999; Olivieri et al. 2006; Plaza et al. 2003; Rando et al. 1998; Stevanovitchet al. 2004; Coudray et al.2008; Cherni et al. 2008
- ↑ Harich et .al 2010, The trans-Saharan slave trade - clues from interpolation analyses and high-resolution characterization of mitochondrial DNA lineages
- ↑ Frigi et. al 2010, Ancient Local Evolution of African mtDNA Haplogroups in Tunisian Berber Populations, Human Biology, Volume 82, Number 4, August 2010
- ↑ Henn, B. M.; Botigué, L. R.; Gravel, S.; Wang, W.; Brisbin, A.; Byrnes, J. K.; Fadhlaoui-Zid, K.; Zalloua, P. A.; Moreno-Estrada, A. (2012). Schierup, Mikkel H, ed. "Genomic Ancestry of North Africans Supports Back-to-Africa Migrations". PLoS Genetics. 8 (1): e1002397. doi:10.1371/journal.pgen.1002397. PMC 3257290. PMID 22253600.
- ↑ Begoña Dobon; Hisham Y. Hassan; Hafid Laayouni; Pierre Luisi; Isis Ricaño-Ponce; Alexandra Zhernakova; Cisca Wijmenga; Hanan Tahir; David Comas; Mihai G. Netea; Jaume Bertranpetit (28 May 2015). "The genetics of East African populations: a Nilo-Saharan component in the African genetic landscape" (PDF). Scientific Reports. 5: 9996. doi:10.1038/srep09996. PMC 4446898. PMID 26017457. Retrieved 18 June 2015.
- ↑ Cruciani et al., 2004, Phylogeography of the Y-Chromosome Haplogroup E3b
- ↑ Reduced Genetic Structure for Iberian Peninsula: implications for population demography. (2004) Archived 2008-04-06 at the Wayback Machine.
- ↑ Goncalves, R.; Freitas, A.; Branco, M.; Rosa, A.; Fernandes, A. T.; Zhivotovsky, L. A.; Underhill, P. A.; Kivisild, T.; Brehm, A. (2005). "Y-chromosome Lineages from Portugal, Madeira and Acores Record Elements of Sephardim and Berber Ancestry". Annals of Human Genetics. 69 (4): 443–54. doi:10.1111/j.1529-8817.2005.00161.x. PMID 15996172.
- ↑ Lacan et al. (2011)
- ↑ "Molecular genetic investigation of the Neolithic population history in the western Carpathian Basin" (PDF).
- ↑ Di Gaetano, C.; Cerutti, N.; Crobu, F.; Robino, C.; Inturri, S.; Gino, S.; Guarrera, S.; Underhill, P. A.; King, R. J.; et al. (2008). "Differential Greek and northern African migrations to Sicily are supported by genetic evidence from the Y chromosome". European Journal of Human Genetics. 17 (1): 91–99. doi:10.1038/ejhg.2008.120. PMC 2985948. PMID 18685561. "The co-occurrence of the Berber E3b1b-M81 (2.12 percent) and of the Mid-Eastern J1-M267 (3.81 percent) Hgs together with the presence of E3b1a1-V12, E3b1a3-V22, E3b1a4-V65 (5.5 percent) support the hypothesis of intrusion of North African genes. (...) These Hgs are common in Northern Africa and are observed only in Mediterranean Europe and together the presence of the E3b1b-M81 highlights the genetic relationships between northern Africa and Sicily. (...) Hg E3b1b-M81 network cluster confirms the genetic affinity between Sicily and North Africa."
- ↑ Adams, S. M.; Bosch, E.; Balaresque, P. L.; Ballereau, S. P. J.; Lee, A. C.; Arroyo, E.; López-Parra, A. M.; Aler, M.; Grifo, M. S. G. (2008). "The Genetic Legacy of Religious Diversity and Intolerance: Paternal Lineages of Christians, Jews, and Muslims in the Iberian Peninsula". The American Journal of Human Genetics. 83 (6): 725–736. doi:10.1016/j.ajhg.2008.11.007. PMC 2668061. PMID 19061982.
- ↑ "The study shows that religious conversions and the subsequent marriages between people of different lineage had in fact a minor impact on modern populations both in Spain, and in Portugal", The religious conversions of Jews and Muslims have affected the population of the Iberian Peninsula Archived 2009-05-21 at the Wayback Machine., Elena Bosch, 2008
- ↑ Plaza, S.; Calafell, F.; Helal, A.; Bouzerna, N.; Lefranc, G.; Bertranpetit, J.; Comas, D. (2003). "Joining the Pillars of Hercules: MtDNA Sequences Show Multidirectional Gene Flow in the Western Mediterranean". Annals of Human Genetics. 67 (4): 312–28. doi:10.1046/j.1469-1809.2003.00039.x. PMID 12914566. But very likely, most of the L mtDNA that has been found in minor amounts in Iberia, is actually pre-neolithic in origin, as it was demonstrated by María Cerezo et al., (Reconstructing ancient mitochondrial DNA links between Africa and Europe). "Haplogroup U6 is present at frequencies ranging from 0-7 percent in the various Iberian populations, with an average of 1.8 percent. Given that the frequency of U6 in NW Africa is 10 percent, the mtDNA contribution of NW Africa to Iberia can be estimated at 18 percent (though U6 has been found in many Iberian hunter-gatherer remains as well). This is larger than the contribution estimated with Y-chromosomal lineages (7 percent) (Bosch et al. 2001).
- ↑ Pereira, L.; Cunha, C.; Alves, C.; Amorim, A. (2005). "African Female Heritage in Iberia: A Reassessment of mtDNA Lineage Distribution in Present Times". Human Biology. 77 (2): 213–229. doi:10.1353/hub.2005.0041. PMID 16201138. "Although the absolute value of observed U6 frequency in Iberia is low, it reveals a discernible North African female contribution, if we keep in mind that haplogroup U6 is not very common in North Africa itself and virtually absent in the rest of Europe. Indeed, because the range of variation in western North Africa is 4-28 percent, the estimated minimum input is 8.54 percent"
- ↑ GonzáLez, A. M.; Brehm, A.; Pérez, J. A.; Maca-Meyer, N.; Flores, C.; Cabrera, V. M. (2003). "Mitochondrial DNA affinities at the Atlantic fringe of Europe". American Journal of Physical Anthropology. 120 (4): 391–404. doi:10.1002/ajpa.10168. PMID 12627534. "Our results clearly reinforce, extend, and clarify the preliminary clues of an 'important very ancient mtDNA contribution from northwest Africa into the Iberian Peninsula' (Côrte-Real et al., 1996; Rando et al., 1998; Flores et al., 2000a; Rocha et al., 1999)(...) Our own data allow us to make minimal estimates of the maternal African pre-Neolithic, Neolithic, and/or recent slave trade input into Iberia. For the former, we consider only the mean value of the U6 frequency in Northern African populations, excluding Saharans, Tuareg, and Mauritanians (16 percent), as the pre-Neolithic frequency in that area, and the present frequency in the whole Iberian Peninsula (2.3 percent) as the result of the northwest African gene flow at that time. The value obtained (14 percent) could be as high as 35 percent using the data of Corte-Real et al. (1996), or 27 percent with our north Portugal sample."
- ↑ Achilli, A.; Olivieri, A.; Pala, M.; Metspalu, E.; Fornarino, S.; Battaglia, V.; Accetturo, M.; Kutuev, I.; Khusnutdinova, E. (2007). "Mitochondrial DNA Variation of Modern Tuscans Supports the Near Eastern Origin of Etruscans". The American Journal of Human Genetics. 80 (4): 759–68. doi:10.1086/512822. PMC 1852723. PMID 17357081. "1.33% (3/226) in Calabria and 1.28 percent in Campania"
- ↑ See the remarks of genetic genealogist Robert Tarín for example. We can add 6.1 percent (eight out of 132) in Cuba
- ↑ Mendizabal, I.; Sandoval, K.; Berniell-Lee, G.; Calafell, F.; Salas, A.; Martinez-Fuentes, A.; Comas, D. (2008). "Genetic origin, admixture, and asymmetry in maternal and paternal human lineages in Cuba". BMC Evolutionary Biology. 8: 213. doi:10.1186/1471-2148-8-213. PMC 2492877. PMID 18644108. "The presence of chromosomes of North African origin (E3b1b-M81; Cruciani et al., 2004) can also be explained by a Portuguese-mediated influx, since this haplogroup reaches a frequency of 5.6 percent in Portugal (Beleza et al., 2006), quite similar to the frequency found in Rio de Janeiro (5.4 percent) among European contributors"
- ↑ Silva, D. A.; Carvalho, E.; Costa, G.; Tavares, L. G.; Amorim, A. N.; Gusmão, L. (2006). "Y-chromosome genetic variation in Rio De Janeiro population". American Journal of Human Biology. 18 (6): 829–837. doi:10.1002/ajhb.20567. PMID 17039481.
- ↑ Coco, C.; Magistrelli, P.; Granone, P.; Roncolini, G.; Picciocchi, A. (1992). "Conservative surgery for early cancer of the distal rectum". Diseases of the Colon & Rectum. 35 (2): 131–136. doi:10.1007/BF02050667.
- ↑ Rabab Khairat; Markus Ball; Chun-Chi Hsieh Chang; Raffaella Bianucci; Andreas G. Nerlich; Martin Trautmann; Somaia Ismail; et al. (4 April 2013). "First insights into the metagenome of Egyptian mummies using next-generation sequencing" (PDF). Journal of Applied Genetics. 54: 309–325. doi:10.1007/s13353-013-0145-1. Retrieved 8 June 2016.
- ↑ Kefi R, Bouzaid E, Stevanovitch A, Beraud-Colomb E. "MITOCHONDRIAL DNA AND PHYLOGENETIC ANALYSIS OF PREHISTORIC NORTH AFRICAN POPULATIONS" (PDF). ISABS. Archived from the original (PDF) on 11 March 2016. Retrieved 17 January 2016.
- ↑ Bernard Secher; Rosa Fregel; José M Larruga; Vicente M Cabrera; Phillip Endicott; José J Pestano; Ana M González. "The history of the North African mitochondrial DNA haplogroup U6 gene flow into the African, Eurasian and American continents". BMC Evolutionary Biology. Retrieved 8 February 2016.
- ↑ Ordóñez, A. C., Fregel, R., Trujillo-Mederos, A., Hervella, M., de-la-Rúa, C., & Arnay-de-la-Rosa, M. (2017). "Genetic studies on the prehispanic population buried in Punta Azul cave (El Hierro, Canary Islands)". Journal of Archaeological Science. 78: 20–28. doi:10.1016/j.jas.2016.11.004. Retrieved 21 April 2017.
- ↑ Fregel et al.2018, Ancient genomes from North Africa evidence prehistoric migrations to the Maghreb from both the Levant and Europe, PNAS June 12, 2018. 201800851; published ahead of print June 12, 2018. https://doi.org/10.1073/pnas.1800851115