Sense (moleculaire biologie)
De sense of polariteit van een nucleïnezuur geeft de richting van het enkelstrengs virale genoom tot de leesrichting van het latere messenger RNA (mRNA) aan. Zoals alle strengen nucleïnezuur heeft mRNA een bepaalde richting. Het ene eind is het 5'-eind (de 5'-cap) en het andere einde het 3'-eind.
Het ribosoom begint bij het vormen van een eiwit met het aflezen (translatie) van het mRNA aan het 5'-eind tot het een startcodon tegenkomt. Vanaf deze plaats worden tijdens het verder aflezen aminozuren aan het eiwit gebonden totdat het ribosoom een stopcodon tegenkomt.
In het algemeen geldt dat een nucleïnezuur, dat in de 5'→3'-afleesrichting van de ribosomen bij de translatie de juiste volgorde van het basentriplet voor het latere eiwit heeft, als sense of als een positieve streng aangeduid wordt.
Bij enkelstrengs (ss, Engels voor single-stranded)-RNA en enkelstrengs-DNA met een positieve polariteit geeft dit de basevolgorde in 5'→3'-richting van het latere mRNA aan. De streng kan namelijk door de ribosomen alleen in de 5'→3'-richting afgelezen worden. Bij een negatieve polariteit is het genoom complementair voor het latere mRNA en kan daardoor niet afgelezen worden. Bij een dubbelstrengs (ds, Engels voor double stranded)-RNA (dsRNA) of -DNA (dsDNA) vindt slechts bij één streng de transcriptie voor het mRNA plaats, omdat de andere streng het spiegelbeeld is en hierdoor niet afgelezen kan worden.
Bij virussen worden drie soorten van polariteit van het genoom onderscheiden:
- sense ((+)-polariteit),
- antisense ((-)-polariteit),
- ambisense: sense en antisense bij dezelfde streng.
Het verschil in polariteit is voor de taxonomische indeling van virussen van groot belang en weerspiegelt de verschillende replicatiestrategieën van de virussen.
Sense

(+)ssRNA-virussen
Bij virussen die een positief enkelstrengs RNA-genoom hebben is de basenvolgorde dezelfde als die van het latere mRNA. Bij deze virussen wordt het (+)ssRNA daarom direct door het ribosoom afgelezen. Alle virussen met een (+)ssRNA-genoom moeten voor een eigen RNA-afhankelijk RNA-polymerase coderen, dat bij de eerste stap van de in de cel binnendringende RNA-streng afgelezen moet worden. De vermeerdering van het virale ssRNA gebeurt met een complementaire (−)-streng als matrijs voor de volgende (+)-strengen. Bekende voorbeelden van dit soort veel voorkomende virussen met (+)ssRNA zijn de Flaviviridae (bijvoorbeeld het hepatitis C-virus) en de Picornaviridae.
(+)ssDNA-virussen
Bij de minder vaak voorkomende (+)ssDNA-virussen gebeurt de synthese van het mRNA via codogenen, een complementaire (-)streng, die alleen tijdens de replicatie in de cel aanwezig is. Daarvoor wordt het enkelstrengs DNA door het in de cel aanwezige DNA-polymerase omgezet in een dubbelstrengs DNA. Een (+)ssDNA komt voor bij de families van de bacteriofagen, de INoviridae en Microviridae (cirkelvormig (+)ssDNA) en bij de plantenvirussen uit de familie Nanoviridae (gesegmenteerd (+)ssDNA).
Antisense

(−)ssRNA-virussen
Bij een (−)ssRNA wordt steeds van het enkelstrengs-RNA een dubbelstrengs-RNA gemaakt. Deze complementaire RNA-streng komt met het mRNA overeen en kan door de ribosomen afgelezen worden. Eukaryotische cellen bezitten geen enzym dat de verdubbeling van het ssRNA tot een dsRNA-streng kan katalyseren. Daarom moeten de virussen met een (−)ssRNA-genoom steeds minstens één molecuul van een van een virus afkomstige RNA-polymerase in hun virusdeeltje ingebouwd hebben, dat dan aan het begin van de virusvermeerdering in de cel de virus eiwit- en RNA-synthese kan opstarten. Virussoorten met (-)ssRNA komen het meeste voor. Hiertoe behoren virussen uit de orde Mononegavirales (een deel van de naam -nega verwijst naar de negativiteit van het ongesegmenteerde genoom) (met bijvoorbeeld de familie Paramyxovirus waartoe het mazelenvirus behoort) en de familie Orthomyxoviridae (met bijvoorbeeld de geslachten van het Influenzavirus).
(−)ssDNA-virussen
Bij (−)ssDNA-virussen wordt het mRNA direct aan deze streng gesynthetiseerd of wordt eerst de streng door in de cel aanwezige DNA-polymerasen verdubbeld. De complementaire DNA-streng komt overeen met de basenvolgorde van het mRNA. Een (−)ssDNA-genoom komt voor bij de Geminiviridae (cirkelvormig (−)ssDNA bij bepaalde plantenvirussen) en bij het geslacht Gyrovirus van de familie Circoviridae (cirkelvormig (−)ssDNA bij vertebratenvirussen van gewervelden).
Ambisense
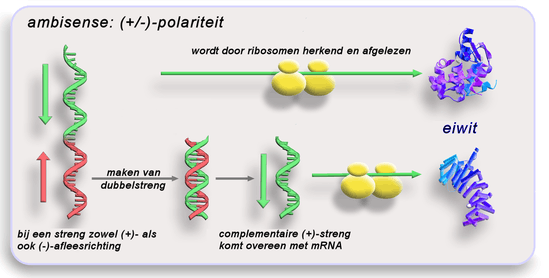
Een bijzondere mengvorm van een sense- en antisense-genoom komt voor bij enkele ssRNA- en ssDNA-virussen. Daarbij komen beide polariteiten op dezelfde streng voor. De genen zijn dan ten minste over twee nucleïnezuurstrengen verdeeld, waarvan eerst een complementaire sequentie gemaakt moet worden. De beide verschillende polariteitsgedeelten worden of onafhankelijk van elkaar eerst in mRNA overgeschreven (in de antisense-gedeelten) dan wel direct als mRNA (in het sense-gedeelte) voor het overeenkomstige mRNA.
Ambisense moet niet verward worden met de gelijktijdige aanwezigheid van zowel enkelstrengs (+) als enkelstrengs (−) in het virusgenoom, zoals bij de Parvoviridae. Dankzij genoomsegmentatie is het immers mogelijk dat een virus het genoom verdeelt in meerdere RNA-moleculen die onafhankelijk van elkaar een eigen polariteit kunnen hebben. Ook moet ambisense niet verward worden met de aanwezigheid van een open leesraam (open reading frame) in beide richtingen van een dubbelstrengs-nucleïnezuur, zoals bij de Polyomaviridae.
Virussen met ambisense-ssRNA zijn de Arenaviridae (bijvoorbeeld Lassavirus, een van de veroorzakers van lassakoorts) en met gedeeltelijk ambisense-ssRNA bij de Bunyaviridae.
Een ambisense-ssDNA komt voor bij het geslacht Circovirus uit de familie Circoviridae.
"Domein" virussen
Afhankelijk van de manier waarop een virus zijn erfelijk materiaal (DNA of RNA) vermenigvuldigt kan ook de volgende kunstmatige indeling in drie groepen worden gemaakt:
- "Domein" virussen (niet-cellulair, plaats onduidelijk, parafyletisch)
- DNA-virussen
- Dubbelstrengs-DNA-virussen (dsDNA, 22 families)
- Enkelstrengs-DNA-virussen (ssDNA, 6 families)
- RNA-virussen
- Dubbelstrengs-RNA-virussen (dsRNA, 7 families)
- Negatief-strengs-enkelstrengs-RNA-virussen (-ssRNA, 7 families)
- Positief-strengs-enkelstrengs-RNA-virussen (+ssRNA, 24 families)
- Retrovirussen
- Subvirale agentia
- DNA-virussen
DNA dubbele helix
Als gevolg van het feit dat de beide strengen van het DNA in tegengestelde richting aan elkaar zitten, en dat het holo-enzym RNA-polymerase voor het aflezen van een stuk DNA de voorkeur heeft voor een bepaalde richting wordt slechts één streng goed afgelezen voor een bepaald gen. De andere streng kan alleen achterstevoren afgelezen worden, en gezien de nodige sequenties voor transcriptie niet aanwezig zijn op die andere streng zal er geen transcriptie optreden. Moleculair biologen noemen een stuk DNA dat afgelezen wordt of afleesbaar is een antisense en het tegenovergestelde, dus niet afleesbaar, sense. Hieruit volgt wat paradoxaal dat de sjabloon (template) voor transcriptie een antisense-streng heeft, hetgeen resulteert in het aflezen van de antisense-DNA-streng in een sense-mRNA-streng.
Bron
- S. J. Flint, L. W. Enquist, V. R. Racaniello und A. M. Skalka: Principles of Virology. Molecular Biology, Pathogenesis, and Control of Animal Viruses. 2. edition, ASM-Press Washington D.C. 2004, ISBN 1-55581-259-7 S. 67ff