Ribosoom
Het ribosoom is een moleculair complex van eiwitten en ribosomaal RNA (rRNA) dat de centrale plaats vormt van eiwitsynthese. Ribosomen komen in grote aantallen voor in het cytoplasma van alle levende cellen. In ribosomen worden aminozuren in een keten aan elkaar verbonden in een volgorde die wordt bepaald door het messenger-RNA (mRNA), een proces genaamd translatie. Ribosomen bestaan uit twee componenten: een klein ribosoomdeel, die het mRNA afleest, en een groot ribosoomdeel, waarin de aminozuurketen wordt gevormd. Ribosomen kunnen vrij voorkomen in het cytoplasma of gebonden zijn aan het endoplasmatisch reticulum of het kernmembraan.
| Celbiologie |
|---|
| De dierlijke cel |
 |
| Portaal |
Cellen besteden veel energie aan de vorming van ribosomen. Een E. coli-bacterie bevat ongeveer 15.000 ribosomen, elk met een molecuulgewicht van ongeveer drie miljoen dalton. Ribosomen vertegenwoordigen circa 25% van de totale massa van deze bacteriën. Ook in planten en dieren is ribosomaal RNA het meest voorkomende type RNA dat in de cel voorkomt.[1]
Algemeen
Genetische informatie die in het DNA vastligt, wordt tijdens de transcriptie overgeschreven naar een mRNA-molecuul. Dit molecuul bevat informatie voor de synthese van eiwitten. Tijdens de translatie wordt de nucleotidevolgorde van het mRNA vertaald naar een aminozuurvolgorde in een polypeptideketen. Hieruit zal een eiwit ontstaan. De translatie speelt zich geheel af in het ribosoom.
Ribosomen herkennen het 5'-eind van de mRNA-keten (de zogenaamde 5'-cap). Het kleine ribosoomdeel bindt aan dit 5'-eind vast en leest de mRNA-keten af richting het 3'-eind totdat een startcodon wordt herkend. Daaraan wordt een methioninecomplex vastgehecht (het eerste aminozuur) en vervolgens bindt ook het grote ribosoomdeel. De translatie kan nu beginnen. Speciale RNA-moleculen binden aminozuren uit het cytoplasma en vervoeren ze naar het ribosoom: transfer-RNA of tRNA. Wanneer tRNA een aminozuur gebonden heeft, gaat het gedeeltelijk het ribosoom binnen en bindt daar tijdelijk aan een complementair gedeelte van de mRNA-keten. Een codon van het mRNA ligt daarbij tegenover een anticodon van het tRNA. Tijdens deze binding wordt het meegevoerde aminozuur met een peptidebinding verbonden aan een al ingebouwd aminozuur. Vervolgens schuift het ribosoom één codon verder langs de mRNA-streng, en kan een nieuw aminozuur worden ingebouwd. Uiteindelijk bereikt het ribosoom een stopcodon, dat voor geen aminozuur codeert. Doordat er geen aminozuur wordt ingebouwd, stopt de eiwitsynthese. De mRNA-keten laat los en de ribosoomdelen gaan uit elkaar.
Ribosomen van bacteriën, archaea en eukaryoten verschillen slechts in detail. Er zijn verschillen in structuur, maar functioneel zijn de ribosomen vrijwel identiek. Dit is een aanwijzing dat ribosomen ontstaan zijn uit een gemeenschappelijke oorsprong. In eukaryotische cellen bevatten mitochondriën een eigen type ribosoom. De mitochondriale ribosomen worden gecodeerd door mitochondriale genen. Deze ribosomen hebben functionele kenmerken die erg lijken op de kenmerken van ribosomen in bacteriën, wat de vermoedelijke evolutionaire oorsprong van mitochondriën aanwijst.[2][3]
Dankzij kristallografisch onderzoek is de kristalstructuur, waarin de honderdduizenden atomen van het ribosoom zijn gerangschikt, inmiddels bekend. In 2011 was de volledige atomaire structuur van een 80S ribosoom vastgesteld. Deze was afkomstig uit de gist Saccharomyces cerevisiae.[4] Daarmee is het ribosoom het grootste niet-virale eiwitcomplex waarvan de structuur is bepaald; ribosomen blijken de molecuulcomplexen te zijn met de meeste asymmetriegerelateerde atomen.
Ontdekking
Ribosomen werden voor het eerst waargenomen door de Roemeens-Amerikaanse celbioloog George Emil Palade in 1955. Hij zag ribosomen met behulp van een elektronenmicroscoop als geclusterde deeltjes of korrels in het cytoplasma.[5] De term 'ribosoom' werd voorgesteld door wetenschapper Richard B. Roberts aan het eind van de jaren vijftig.
Albert Claude, Christian de Duve en George Emil Palade kregen in 1974 gezamenlijk de Nobelprijs voor Fysiologie of Geneeskunde voor de ontdekking van het ribosoom.[6] De Nobelprijs voor de Scheikunde 2009 werd uitgereikt aan Venkatraman Ramakrishnan, Thomas Steitz en Ada Yonath voor het gedetailleerd vaststellen van de structuur en werking van het ribosoom.[7]
Structuur
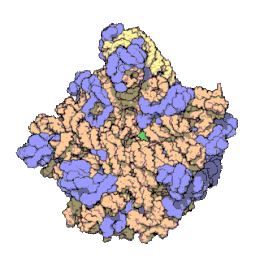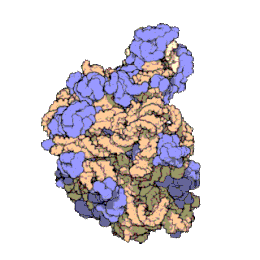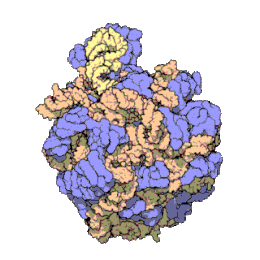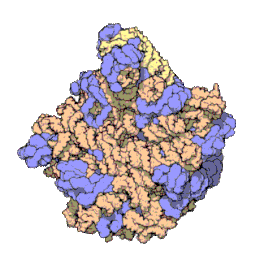
Het ribosoom is een zeer complex moleculair systeem. In vergelijking met andere organellen in de cel zijn ribosomen zeer kleine structuren. Een ribosoom bestaat grotendeels uit een gespecialiseerd type RNA genaamd ribosomaal RNA (rRNA) en een groot aantal verschillende ribosomale eiwitten, waarvan het exacte aantal tussen soorten varieert. De ribosomale eiwitten (r-eiwitten) en rRNA's vouwen om elkaar heen en vormen dan twee afzonderlijke ribosomale delen: een groot en een klein ribosoomdeel. De twee delen passen in elkaar en werken samen als één om eiwitsynthese te faciliteren. Omdat ribosomen uit twee subunits bestaan van ongelijke grootte, zijn ze iets langer in de hoogte dan in breedte.
Prokaryotische ribosomen hebben een diameter van ongeveer 20 nm (200 Å) en zijn opgebouwd uit 65% rRNA- en 35% ribosomale eiwitten.[9] Eukaryotische ribosomen hebben een diameter tussen 25 en 30 nm (250-300 Å) en bestaan ongeveer voor de helft uit rRNA en voor de andere helft uit ribosomale eiwitten.[10] Kristallografisch onderzoek heeft aangetoond dat er geen ribosomale eiwitten voorkomen in het actieve centrum van ribosomen.[11] Dit suggereert dat de eiwitcomponenten niet direct deelnemen aan het katalyseren van de vorming van peptidebindingen, maar dat deze eiwitten zorgen voor stabilisering van de correcte structuur die het katalytische vermogen van rRNA-ketens optimaliseren. De eiwitten, die voornamelijk aan de buitenkant voorkomen, ondersteunen ook de vormveranderingen van de rRNA-ketens die optreden tijdens translatie.
De eenheid die wordt gebruikt om het ribosoom en de rRNA-ketens te beschrijven, is de Svedberg-eenheid (S). Het is een eenheid voor de sedimentatiesnelheid van moleculen en wordt bepaald door de grootte, massa en vorm van een molecuul. De sedimentatiesnelheden van de twee aparte ribosoomdelen kunnen opgeteld groter zijn dan die van het gehele ribosoom: prokaryotische 70S-ribosomen zijn bijvoorbeeld gemaakt van 50S- en 30S-ribosoomdelen.
Prokaryoten
Ribosomen worden gevormd wanneer rRNA-genen worden afgeschreven en de ontstane rRNA-ketens zich met r-eiwitten in een juiste ruimtelijke structuur organiseren. Prokaryoten hebben 70S-ribosomen, elk bestaande uit een klein (30S) en een groot (50S) ribosoomdeel. Het kleine ribosoomdeel bevat een 16S rRNA-keten (bestaande uit 1540 nucleotiden) en bestaat uit 21 eiwitten. Het grote ribosoomdeel bestaat uit een 5S-RNA-keten (120 nucleotiden), een 23S-RNA-keten (2900 nucleotiden) en 31 eiwitten. Zie onderstaand schema.
Eukaryoten
In eukaryoten worden de ribosoomdelen gevormd in de nucleolus. Via kernporiën worden ze naar het cytoplasma vervoert. Eukaryotische cellen hebben 80S-ribosomen, wederom bestaande uit een klein (40S) en groot (60S) ribosoomdeel. Het 40S-deel bevat een 18S rRNA-keten (1900 nucleotiden) en 33 eiwitten. Het grote ribosoomdeel bestaat uit een 5S-RNA (120 nucleotiden), 28S-RNA (4700 nucleotiden), 5,8S-RNA (160 nucleotiden) en 46 eiwitten. Zie onderstaand schema.
eukaryotisch vrij ribosoom (R. norvegicus)[12] ribosoom ribosoomdeel rRNA-ketens r-eiwitten 80S 60S 28S (4718 nt) 49 5,8S (160 nt) 5S (120 nt) 40S 18S (1874 nt) 33
De ribosomen uit chloroplasten en mitochondriën bestaan ook uit grote en kleine ribosoomdelen die samen met eiwitten zijn verbonden tot een 70S-complex. Volgens de embosymbiosetheorie zouden deze ribosomen ontstaan zijn uit bacteriën. Mitochondriale ribosomen zijn dan ook zeer vergelijkbaar met die van bacteriën.
Het feit dat bacteriële en eukaryotische ribosomen in enige mate verschillen is medisch van belang. Het antibioticum chlooramfenicol werkt, door de verschillen in structuur, wel in op bacteriële, maar niet op eukaryotische ribosomen. Een bacteriële infectie kan worden bestreden doordat de eiwitsynthese van de ziekteverwekkende bacterie wordt geïnhibeerd, terwijl de menselijke cellen onaangetast blijven. Ook tetracycline en streptomycine worden gebruikt om op deze manier een bacteriële infectie tegen te gaan. Hoewel mitochondriën ribosomen hebben die lijken op de bacteriële ribosomen, treedt in mitochondriën geen inhibitie op door deze antibiotica omdat ze zijn omgeven door een dubbel membraan waar antibiotica moeilijk door kunnen passeren.[13]
Actieve centrum
Het actieve centrum van ribosomen wordt, in tegenstelling tot die van enzymen, volledig gevormd door de rRNA-ketens. In het actieve centrum vindt de katalyse van peptidebindingen plaats tussen de aminozuren. Deze plek wordt daarom ook wel het peptidyltransferasecentrum genoemd. Ribosomen werken als katalysatoren voor peptidyltransferase (vorming van een peptidebinding tussen aminozuren) en peptidylhydrolyse (het loskoppelen van aminozuurketen en de tRNA). Het peptidyltransferase-centrum is verantwoordelijk voor het produceren van polypeptiden tijdens de eiwitelongatie. Ribosomen kunnen door hun katalytische activiteit beschouwd worden als grote ribozymen.
Functie
Ribosomen zijn betrokken bij het synthetiseren van alle eiwitten. Ze hebben de taak om de tRNA-moleculen op de juiste manier te binden aan de codons in het mRNA zodat een aminozuur op de goede plek in de keten wordt ingebouwd. Eiwitten regelen de stofwisseling, het transport en de communicatie van cellen in alle levensvormen. Direct en indirect zijn ze betrokken bij alle levensprocessen.
De structuur van ribosomen weerspiegelt hun functie om mRNA en tRNA-dragende aminozuren bij elkaar te brengen. Wanneer het mRNA wordt afgelezen, passeert elk codon achtereenvolgens de A-plaats, de P-plaats en de E-plaats in het ribosoom. Op de A-plaats (aminoacyl-binding-site) bindt het tRNA tijdelijk aan de mRNA-streng doordat het anticodon en het codon waterstofbruggen vormen. Op de P-plaats (peptidyl-binding-site) wordt de aminozuurketen gevormd doordat er een peptidebinding ontstaat tussen het aminozuur in de P-plaats en de A-plaats. Deze plaats is het actieve centrum (het peptidyltransferase centrum), en is vrijwel volledig opgebouwd uit RNA. Op de E-plaats (exit-site) verlaat tRNA zonder aminozuur het ribosoom.
Eiwitsynthese begint bij een startcodon AUG nabij het 5'-eind van het mRNA. Het ribosoom weet waar aan de mRNA-keten het zich moet binden doordat het een specifieke nucleotidevolgorde herkent: de Shine-Dalgarnosequentie in prokaryoten en de Kozak-sequentie in eukaryoten. Aan het eind van de mRNA-keten zit een stopcodon, waaraan geen tRNA kan hechten. Doordat geen aminozuur wordt ingebouwd, stopt de eiwitsynthese. De aminozuurketen laat los en het kleine en grote ribosoomdeel gaan uit elkaar.
Er kunnen meerdere ribosomen tegelijkertijd gebonden zijn aan één mRNA-molecuul. Zo kan veel van hetzelfde eiwit in de cel worden geproduceerd. Met een elektronenmicroscoop zijn clusters van ribosomen (polyribosomen) in de cel zichtbaar.
Binding aan ER
Ribosomen komen vrij voor in het cytoplasma of gebonden aan het endoplasmatisch reticulum (ER) of kernmembraan. De eiwitten die worden gemaakt in vrije ribosomen functioneren meestal binnen het cytosol, bijvoorbeeld een enzym dat een stap in de glycolyse katalyseert. Ribosomen aan het endoplasmatisch reticulum zijn betrokken bij de synthese van membraaneiwitten, eiwitten die verpakt moeten worden (zoals lysozymen) en bij eiwitten die aan het externe milieu van de cel worden afgegeven (exocytose). Cellen die zich specialiseren in eiwitsecretie, zoals alvleeskliercellen die spijsverteringsenzymen afscheiden, hebben vaak zeer veel gebonden ribosomen. Er is geen verschil in structuur tussen gebonden en vrije ribosomen; of het ribosoom al dan niet aan het ER bindt hangt af van welk eiwit het op dat moment synthetiseert.
De ribosoomdelen, afkomstig uit de nucleolus, omsluiten een mRNA-keten altijd in het cytosol. Als het om de synthese van een secretie-eiwit gaat, moet het ribosoom zich hechten aan het ER-membraan. Aan het uiteinde van de groeiende peptideketen bij secretie-eiwitten komt een specifieke sequentie voor die door zogenaamde SRP-eiwitten wordt herkend.[14] Het SRP-eiwit bindt in dat geval aan de polypeptideketen (en daarbij ook aan het ribosoom) en de translatie wordt kort onderbroken: tRNA-moleculen komen niet meer naar binnen. Het SRP-gebonden ribosoom kan nu op een specifieke plek aan het ER-membraan hechten, namelijk op een SRP-receptorplaats die naast een translocon (kanaaltje voor peptiden) is gelegen.[15] Het ribosoom wordt zo gebonden aan het translocon, waardoorheen het polypeptide gesynthetiseerd kan worden. Het SRP-eiwit laat los en de translatie wordt voortgezet.
Oorsprong
Ribosomen zijn waarschijnlijk verschenen in de RNA-wereld als zelfreplicerende structuurtjes.[16] Aannemelijk is dat deze vroege ribosomen nog uitsluitend uit rRNA waren opgebouwd. Later, toen interacties met individuele aminozuren waren ontstaan, ontwikkelden ribosomen geleidelijk het vermogen om peptidebindingen te katalyseren.[17][18] Onderzoek wijst uit dat in de eerste rRNA-moleculen van ribosomen ook coderende stukken aanwezig waren. De rRNA-ketens codeerden voor tRNA-moleculen die het ribosoom gebruikte voor zelfreplicatie.[19]
Naarmate aminozuren in de RNA-wereld verschenen, konden ze door hun interacties met RNA de efficiëntie vergroten van de functie van katalytische RNA-moleculen. De drijvende kracht achter de evolutie van het ribosoom, van een primitieve zelfreplicerende structuur naar zijn huidige vorm, kan de selectiedruk zijn geweest om eiwitten in het ribosoom op te nemen, om zo zijn eigen zelfreplicatie te garanderen.[19]
Zie ook
Referenties
Literatuur
|
| Celorganellen van eukaryoten | ||||||
|---|---|---|---|---|---|---|
|