Settlement of the Americas

The first settlement of the Americas began when Paleolithic hunter-gatherers first entered North America from the North Asian Mammoth steppe via the Beringia land bridge, which had formed between northeastern Siberia and western Alaska due to the lowering of sea level during the Last Glacial Maximum. These populations expanded south of the Laurentide Ice Sheet and rapidly throughout both North and South America, by 14,000 years ago.[1][2][3][4] The earliest populations in the Americas, before roughly 10,000 years ago, are known as Paleo-Indians.
The peopling of the Americas was a long-standing open question, and while advances in archaeology, Pleistocene geology, physical anthropology, and DNA analysis have shed progressively more light on the subject, significant questions remain unresolved.[5] While there is general agreement that the Americas were first settled from Asia, the pattern of migration, its timing, and the place(s) of origin in Eurasia of the peoples who migrated to the Americas remain unclear.[2]
The prevalent migration models outline different time frames for the Asian migration from the Bering Straits and subsequent dispersal of the founding population throughout the continent.[6] Indigenous peoples of the Americas have been linked to Siberian populations by linguistic factors, the distribution of blood types, and in genetic composition as reflected by molecular data, such as DNA.[7]
The "Clovis first theory" refers to the 1950s hypothesis that the Clovis culture represents the earliest human presence in the Americas, beginning about 13,000 years ago; evidence of pre-Clovis cultures has accumulated since 2000, pushing back the possible date of the first peopling of the Americas to about 13,200–15,500 years ago.[8][9]
The environment during the latest Pleistocene
For an introduction to the radiocarbon dating techniques used by archaeologists and geologists, see radiocarbon dating.
Emergence and submergence of Beringia
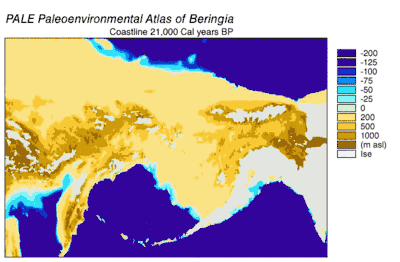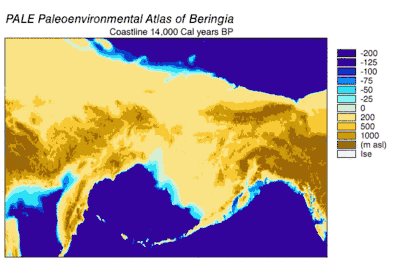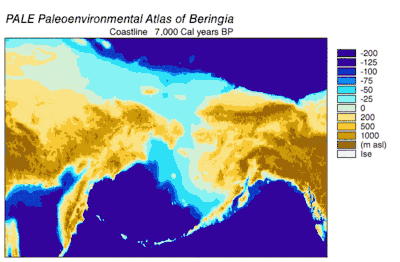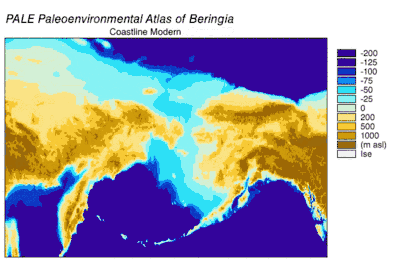
During the Wisconsin Glaciation, varying portions of the Earth's water were stored as glacier ice. As water accumulated in glaciers, the volume of water in the oceans correspondingly decreased, resulting in lowering of global sea level. The variation of sea level over time has been reconstructed using oxygen isotope analysis of deep sea cores, the dating of marine terraces, and high resolution oxygen isotope sampling from ocean basins and modern ice caps. A drop of eustatic sea level by about 60 m to 120 m lower than present-day levels, commencing around 30,000 years BP, created Beringia, a durable and extensive geographic feature connecting Siberia with Alaska.[10] With the rise of sea level after the Last Glacial Maximum (LGM), the Beringian land bridge was again submerged. Estimates of the final re-submergence of the Beringian land bridge based purely on present bathymetry of the Bering Strait and eustatic sea level curve place the event around 11,000 years BP (Figure 1). Ongoing research reconstructing Beringian paleogeography during deglaciation could change that estimate and possible earlier submergence could further constrain models of human migration into North America.[10]
Glaciers
The onset of the Last Glacial Maximum after 30,000 years BP saw the expansion of alpine glaciers and continental ice sheets that blocked migration routes out of Beringia. By 21,000 years BP, and possibly thousands of years earlier, the Cordilleran and Laurentide ice sheets coalesced east of the Rocky Mountains, closing off a potential migration route into the center of North America.[11][12][13] Alpine glaciers in the coastal ranges and the Alaskan Peninsula isolated the interior of Beringia from the Pacific coast. Coastal alpine glaciers and lobes of Cordilleran ice coalesced into piedmont glaciers that covered large stretches of the coastline as far south as Vancouver Island and formed an ice lobe across the Straits of Juan de Fuca by 15,000 14C years BP (18,000 cal years BP).[14][15] Coastal alpine glaciers started to retreat around 19,000 cal years BP [16] while Cordilleran ice continued advancing in the Puget lowlands up to 14,000 14C years BP (16,800 cal years BP)[15] Even during the maximum extent of coastal ice, unglaciated refugia persisted on present-day islands, that supported terrestrial and marine mammals.[13] As deglaciation occurred, refugia expanded until the coast became ice-free by 15,000 cal years BP.[13] The retreat of glaciers on the Alaskan Peninsula provided access from Beringia to the Pacific coast by around 17,000 cal years BP.[17] The ice barrier between interior Alaska and the Pacific coast broke up starting around 13,500 14C years (16,200 cal years) BP.[14] The ice-free corridor to the interior of North America opened between 13,000 and 12,000 cal years BP.[11][12][13] Glaciation in eastern Siberia during the LGM was limited to alpine and valley glaciers in mountain ranges and did not block access between Siberia and Beringia.[10]
Climate and biological environments
The paleoclimates and vegetation of eastern Siberia and Alaska during the Wisconsin glaciation have been deduced from high resolution oxygen isotope data and pollen stratigraphy.[10][18][19] Prior to the Last Glacial Maximum, climates in eastern Siberia fluctuated between conditions approximating present day conditions and colder periods. The pre-LGM warm cycles in Arctic Siberia saw flourishes of megafaunas.[10] The oxygen isotope record from the Greenland Ice Cap suggests that these cycles after about 45k years BP lasted anywhere from hundreds to between one and two thousand years, with greater duration of cold periods starting around 32k cal years BP.[10] The pollen record from Elikchan Lake, north of the Sea of Okhotsk, shows a marked shift from tree and shrub pollen to herb pollen prior to 26k 14C years BP, as herb tundra replaced boreal forest and shrub steppe going into the LGM.[10] A similar record of tree/shrub pollen being replaced with herb pollen as the LGM approached was recovered near the Kolyma River in Arctic Siberia.[19] The abandonment of the northern regions of Siberia due to rapid cooling or the retreat of game species with the onset of the LGM has been proposed to explain the lack of archaeosites in that region dating to the LGM.[19][20] The pollen record from the Alaskan side shows shifts between herb/shrub and shrub tundra prior to the LGM, suggesting less dramatic warming episodes than those that allowed forest colonization on the Siberian side. Diverse, though not necessarily plentiful, megafaunas were present in those environments. Herb tundra dominated during the LGM, due to cold and dry conditions.[18]
Coastal environments during the Last Glacial Maximum were complex. The lowered sea level, and an isostatic bulge equilibrated with the depression beneath the Cordilleran Ice Sheet, exposed the continental shelf to form a coastal plain.[21] While much of the coastal plain was covered with piedmont glaciers, unglaciated refugia supporting terrestrial mammals have been identified on Haida Gwaii, Prince of Wales Island, and outer islands of the Alexander Archipelago.[18] The now-submerged coastal plain has potential for more refugia.[18] Pollen data indicate mostly herb/shrub tundra vegetation in unglaciated areas, with some boreal forest towards the southern end of the range of Cordilleran ice.[18] The coastal marine environment remained productive, as indicated by fossils of pinnipeds.[21] The highly productive kelp forests over rocky marine shallows may have been a lure for coastal migration.[22][23] Reconstruction of the southern Beringian coastline also suggests potential for a highly productive coastal marine environment.[23]
Environmental changes during deglaciation
Pollen data indicate a warm period culminating between 14k and 11k 14C years BP (17k-13k cal years BP) followed by cooling between 11k-10k 14C years BP (13k-11.5k cal years BP).[21] Coastal areas deglaciated rapidly as coastal alpine glaciers, then lobes of Cordilleran ice, retreated. The retreat was accelerated as sea levels rose and floated glacial termini. Estimates of a fully ice-free coast range between 16k[21] and 15k[13] cal years BP. Littoral marine organisms colonized shorelines as ocean water replaced glacial meltwater. Replacement of herb/shrub tundra by coniferous forests was underway by 12.4k 14C years BP (15k cal years BP) north of Haida Gwaii. Eustatic sea level rise caused flooding, which accelerated as the rate grew more rapid.[21]
The inland Cordilleran and Laurentide ice sheets retreated more slowly than did the coastal glaciers. Opening of an ice-free corridor did not occur until after 13k to 12k cal years BP.[11][12][13] The early environment of the ice-free corridor was dominated by glacial outwash and meltwater, with ice-dammed lakes and periodic flooding from the release of ice-dammed meltwater.[11] Biological productivity of the deglaciated landscape was gained slowly.[13] The earliest possible viability of the ice-free corridor as a human migration route has been estimated at 11.5k cal years BP.[13]
Birch forests were advancing across former herb tundra in Beringia by 14.3ka 14C years BP (17k cal years BP) in response to climatic amelioration, indicating increased productivity of the landscape.[19]
Chronology and sources of migration
The archaeological community is in general agreement that the ancestors of the Indigenous peoples of the Americas of historical record entered the Americas at the end of the Last Glacial Maximum (LGM), shortly after 20,000 years ago, with ascertained archaeological presence shortly after 16,000 years ago.
There remain uncertainties regarding the precise dating of individual sites and regarding conclusions drawn from population genetics studies of contemporary Native Americans. It is also an open question whether this post-LGM migration represented the first peopling of the Americas, or whether there had been an earlier, pre-LGM migration which had reached South America as early as 40,000 years ago.
Chronology
In the early 21st century, the models of the chronology of migration are divided into two general approaches.[24][25]
The first is the short chronology theory, that the first migration occurred after the Last Glacial Maximum, which went into decline after about 19,000 years ago,[16] and was then followed by successive waves of immigrants.[26]
The second theory is the long chronology theory, which proposes that the first group of people entered the Americas at a much earlier date, possibly before 40,000 years ago,[27][28][29] followed by a much later second wave of immigrants.[25][30]
The Clovis First theory, which dominated thinking on New World anthropology for much of the 20th century, was challenged by the secure dating of archaeosites in the Americas to before 13kya in the 2000s.[31][32][11][12][13] The "short chronology" scenario, in the light of this, refers to a peopling of the Americas shortly after 19,000 years ago, while the "long chronology" scenario permits pre-LGM presence, by around 40 kya.
Evidence for pre-LGM human presence
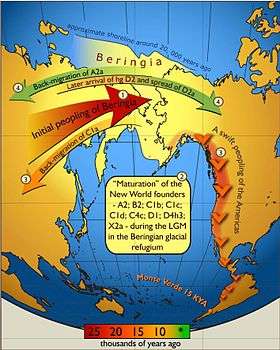
!["Maps depicting each phase of the three-step early human migrations for the peopling of the Americas. (A) Gradual population expansion of the Amerind ancestors from their East Central Asian gene pool (blue arrow). (B) Proto-Amerind occupation of Beringia with little to no population growth for ≈20,000 years. (C) Rapid colonization of the New World by a founder group migrating southward through the ice-free, inland corridor between the eastern Laurentide and western Cordilleran Ice Sheets (green arrow) and/or along the Pacific coast (red arrow). In (B), the exposed seafloor is shown at its greatest extent during the last glacial maximum at ≈20–18 kya [25]. In (A) and (C), the exposed seafloor is depicted at ≈40 kya and ≈16 kya, when prehistoric sea levels were comparable. A scaled-down version of Beringia today (60% reduction of A–C) is presented in the lower left corner. This smaller map highlights the Bering Strait that has geographically separated the New World from Asia since ≈11–10 kya."](../I/m/Journal.pone.0001596.g004.png)
Pre-Last Glacial Maximum migration across Beringia into the Americas has been proposed to explain purported pre-LGM ages of archaeosites in the Americas such as Bluefish Caves[28] and Old Crow Flats[29] in the Yukon Territory, and Meadowcroft Rock Shelter in Pennsylvania.[25][30] The earlier 14C date from a bone artifact at the Old Crow Flats site has been supplanted by an Accelerator Mass Spectrometry 14C date that indicates a Holocene age.[34]
Pre-LGM human presence in South America rests partly on the chronology of the controversial Pedra Furada rock shelter in Piauí, Brazil. A 2003 study dated evidence for the controlled use of fire to before 40 kya.[35] Additional evidence has been adduced from the morphology of Luzia Woman fossil, which was described as Australoid. This interpretation was challenged in a 2003 review which concluded the features in question could also have arisen by genetic drift.[36]
The interpretations of butcher marks and the geologic association of bones at the Bluefish Cave and Old Crow Flats sites have been called into question.[37] The ages of the earliest positively identified artifacts at the Meadowcroft site are constrained by a compiled age estimate from 14C in the range of 12k-15k 14C years BP (13.8k-18.5k cal years BP).[31][38] The Meadowcroft Rockshelter site and the Monte Verde site in southern Chile, with a date of 14.8k cal years BP,[32] are the archaeosites in the Americas with the oldest dates that have gained broad acceptance.
Stones described as probable tools, hammerstones and anvils, have been found in southern California, at the Cerutti Mastodon site, that are associated with a mastodon skeleton which appeared to have been processed by humans. The mastodon skeleton was dated by thorium-230/uranium radiometric analysis, using diffusion–adsorption–decay dating models, to 130.7 ± 9.4 thousand years ago.[39] No human bones were found, and the claims of tools and bone processing have been described as "not plausible".[40]
The Yana River Rhino Horn site (RHS) has dated human occupation of eastern Arctic Siberia to 27k 14C years BP (31.3k cal years BP).[41] That date has been interpreted by some as evidence that migration into Beringia was imminent, lending credence to occupation of Beringia during the LGM.[42][43] However, the Yana RHS date is from the beginning of the cooling period that led into the LGM.[10] But, a compilation of archaeosite dates throughout eastern Siberia suggest that the cooling period caused a retreat of humans southwards.[19][20] Pre-LGM lithic evidence in Siberia indicate a settled lifestyle that was based on local resources, while post-LGM lithic evidence indicate a more migratory lifestyle.[20]
The oldest archaeosite on the Alaskan side of Beringia date to 12k 14C years BP (14k cal years BP).[19][34] It is possible that a small founder population had entered Beringia before that time. However, archaeosites that date closer to the Last Glacial Maximum on either the Siberian or the Alaskan side of Beringia are lacking.
Genomic age estimates
Studies of Amerindian genetics have used high resolution analytical techniques applied to DNA samples from modern Native Americans and Asian populations regarded as their source populations to reconstruct the development of human Y-chromosome DNA haplogroups (yDNA haplogroups) and human mitochondrial DNA haplogroups (mtDNA haplogroups) characteristic of Native American populations.[27][42][43] Models of molecular evolution rates were used to estimate the ages at which Native American DNA lineages branched off from their parent lineages in Asia and to deduce the ages of demographic events. One model (Tammetal 2007) based on Native American mtDNA Haplotypes (Figure 2) proposes that migration into Beringia occurred between 30k and 25k cal years BP, with migration into the Americas occurring around 10k to 15k years after isolation of the small founding population.[42] Another model (Kitchen et al. 2008) proposes that migration into Beringia occurred approximately 36k cal years BP, followed by 20k years of isolation in Beringia.[43] A third model (Nomatto et al. 2009) proposes that migration into Beringia occurred between 40k and 30k cal years BP, with a pre-LGM migration into the Americas followed by isolation of the northern population following closure of the ice-free corridor.[27] Evidence of Australo-Melanesians admixture in Amazonian populations was found by Skoglund and Reich (2016).[44]
A study of the diversification of mtDNA Haplogroups C and D from southern Siberia and eastern Asia, respectively, suggests that the parent lineage (Subhaplogroup D4h) of Subhaplogroup D4h3, a lineage found among Native Americans and Han Chinese,[45][46] emerged around 20k cal years BP, constraining the emergence of D4h3 to post-LGM.[47] Age estimates based on Y-chromosome micro-satellite diversity place origin of the American Haplogroup Q1a3a (Y-DNA) at around 10k to 15k cal years BP.[48] Greater consistency of DNA molecular evolution rate models with each other and with archaeological data may be gained by the use of dated fossil DNA to calibrate molecular evolution rates.[45]
Source populations
There is general agreement among anthropologists that the source populations for the migration into the Americas originated from an area somewhere east of the Yenisei River. The common occurrence of the mtDNA Haplogroups A, B, C, and D among eastern Asian and Native American populations has long been recognized, along with the presence of Haplogroup X.[49] As a whole, the greatest frequency of the four Native American associated haplogroups occurs in the Altai-Baikal region of southern Siberia.[50] Some subclades of C and D closer to the Native American subclades occur among Mongolian, Amur, Japanese, Korean, and Ainu populations.[49][51]
Human genomic models
The development of high-resolution genomic analysis has provided opportunities to further define Native American subclades and narrow the range of Asian subclades that may be parent or sister subclades. For example, the broad geographic range of Haplogroup X has been interpreted as allowing the possibility of a western Eurasian, or even a European source population for Native Americans, as in the Solutrean hypothesis, or suggesting a pre-Last Glacial Maximum migration into the Americas.[49] The analysis of an ancient variant of Haplogroup X among aboriginals of the Altai region indicates common ancestry with the European strain rather than descent from the European strain.[50] Further division of X subclades has allowed identification of Subhaplogroup X2a, which is regarded as specific to Native Americans.[42][46] With further definition of subclades related to Native American populations, the requirements for sampling Asian populations to find the most closely related subclades grow more specific. Subhaplogroups D1 and D4h3 have been regarded as Native American specific based on their absence among a large sampling of populations regarded as potential descendants of source populations, over a wide area of Asia.[42] Among the 3764 samples, the Sakhalin - lower Amur region was represented by 61 Oroks.[42] In another study, Subhaplogroup D1a has been identified among the Ulchis of the lower Amur River region(4 among 87 sampled, or 4.6%), along with Subhaplogroup C1a (1 among 87, or 1.1%).[51] Subhaplogroup C1a is regarded as a close sister clade of the Native American Subhaplogroup C1b.[51] Subhaplogroup D1a has also been found among ancient Jōmon skeletons from Hokkaido[52] The modern Ainu are regarded as descendants of the Jōmon.[52] The occurrence of the Subhaplogroups D1a and C1a in the lower Amur region suggests a source population from that region distinct from the Altai-Baikal source populations, where sampling did not reveal those two particular subclades.[51] The conclusions regarding Subhaplogroup D1 indicating potential source populations in the lower Amur[51] and Hokkaido[52] areas stand in contrast to the single-source migration model.[27][42][43]
Subhaplogroup D4h3 has been identified among Han Chinese.[45][46] Subhaplogroup D4h3 from China does not have the same geographic implication as Subhaplotype D1a from Amur-Hokkaido, so its implications for source models are more speculative. Its parent lineage, Subhaplotype D4h, is believed to have emerged in east Asia, rather than Siberia, around 20k cal years BP.[47] Subhaplogroup D4h2, a sister clade of D4h3, has also been found among Jōmon skeletons from Hokkaido.[53] D4h3 has a coastal trace in the Americas.[46]
The contrast between the genetic profiles of the Hokkaido Jōmon skeletons and the modern Ainu illustrates another uncertainty in source models derived from modern DNA samples:[52]
However, probably due to the small sample size or close consanguinity among the members of the site, the frequencies of the haplogroups in Funadomari skeletons were quite different from any modern populations, including Hokkaido Ainu, who have been regarded as the direct descendant of the Hokkaido Jomon people.
The descendants of source populations with the closest relationship to the genetic profile from the time when differentiation occurred are not obvious. Source population models can be expected to become more robust as more results are compiled, the heritage of modern proxy candidates becomes better understood, and fossil DNA in the regions of interest is found and considered.
HTLV-1 genomics
The Human T cell Lymphotrophic Virus 1 (HTLV-1) is a virus transmitted through exchange of bodily fluids and from mother to child through breast milk. The mother-to-child transmission mimics a hereditary trait, although such transmission from maternal carriers is less than 100%.[54] The HTLV virus genome has been mapped, allowing identification of four major strains and analysis of their antiquity through mutations. The highest geographic concentrations of the strain HLTV-1 are in sub-Saharan Africa and Japan.[55] In Japan, it occurs in its highest concentration on Kyushu.[55] It is also present among African descendants and native populations in the Caribbean region and South America.[55] It is rare in Central America and North America.[55] Its distribution in the Americas has been regarded as due to importation with the slave trade.[56]
The Ainu have developed antibodies to HTLV-1, indicating its endemicity to the Ainu and its antiquity in Japan.[57] A subtype "A" has been defined and identified among the Japanese (including Ainu), and among Caribbean and South American isolates.[58] A subtype "B" has been identified in Japan and India.[58] In 1995, Native Americans in coastal British Columbia were found to have both subtypes A and B.[59] Bone marrow specimens from an Andean mummy about 1500 years old were reported to have shown the presence of the A subtype.[60] The finding ignited controversy, with contention that the sample DNA was insufficiently complete for the conclusion and that the result reflected modern contamination.[61] However, a re-analysis indicated that the DNA sequences were consistent with, but not definitely from, the "cosmopolitan clade" (subtype A).[61] The presence of subtypes A and B in the Americas is suggestive of a Native American source population related to the Ainu ancestors, the Jōmon.
Physical anthropology
Paleoamerican skeletons in the Americas such as Kennewick Man (Washington State), Hoya Negro skeleton (Yucatán), Luzia Woman and other skulls from the Lagoa Santa site (Brazil), Buhl Woman (Idaho), Peñon Woman III,[62] two skulls from the Tlapacoya site (Mexico City),[62] and 33 skulls from Baja California[63] have exhibited craniofacial traits distinct from most modern Native Americans, leading physical anthropologists to the opinion that some Paleoamericans were of an Australoid rather than Siberian origin.[64] The most basic measured distinguishing trait is the dolichocephaly of the skull. Some modern isolates such as the Pericúes of Baja California and the Fuegians of Tierra del Fuego exhibit that same morphological trait.[63] Other anthropologists advocate an alternative hypothesis that evolution of an original Beringian phenotype gave rise to a distinct morphology that was similar in all known Paleoamerican skulls, followed by later convergence towards the modern Native American phenotype.[65][66] Resolution of the issue awaits the identification of a Beringian phenotype among paleoamerican skulls or evidence of a genetic clustering among examples of the Australoid phenotype.
A report published in the American Journal of Physical Anthropology in January 2015 reviewed craniofacial variation focussing on differences between early and late Native Americans and explanations for these based on either skull morphology or molecular genetics. Arguments based on molecular genetics have in the main, according to the authors, accepted a single migration from Asia with a probable pause in Berengia, plus later bi-directional gene flow. Studies focussing on craniofacial morphology have argued that Paleoamerican remains have "been described as much closer to African and Australo-Melanesians populations than to the modern series of Native Americans", suggesting two entries into the Americas, an early one occurring before a distinctive East Asian morphology developed (referred to in the paper as the "Two Components Model". A third model, the "Recurrent Gene Flow" [RGF] model, attempts to reconcile the two, arguing that circumarctic gene flow after the initial migration could account for morphological changes. It specifically re-evaluates the original report on the Hoya Negro skeleton which supported the RGF model, the authors disagreed with the original conclusion which suggested that the skull shape did not match those of modern Native Americans, arguing that the "skull falls into a subregion of the morphospace occupied by both Paleoamericans and some modern Native Americans."[67][68]
Stemmed points
Stemmed points are a lithic technology distinct from Beringian and Clovis types. They have a distribution ranging from coastal east Asia to the Pacific coast of South America.[22] The emergence of stemmed points has been traced to Korea during the upper Paleolithic.[69] The origin and distribution of stemmed points have been interpreted as a cultural marker related to a source population from coastal east Asia.[22]
Migration routes
Interior route
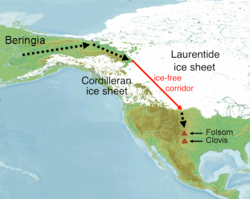
Historically, theories about migration into the Americas have centered on migration from Beringia through the interior of North America. The discovery of artifacts in association with Pleistocene faunal remains near Clovis, New Mexico in the early 1930s required extension of the timeframe for the settlement of North America to the period during which glaciers were still extensive. That led to the hypothesis of a migration route between the Laurentide and Cordilleran ice sheets to explain the early settlement. The Clovis site was host to a lithic technology characterized by spear points with an indentation, or flute, where the point was attached to the shaft. A lithic complex characterized by the Clovis Point technology was subsequently identified over much of North America and in South America. The association of Clovis complex technology with late Pleistocene faunal remains led to the theory that it marked the arrival of big game hunters that migrated out of Beringia then dispersed throughout the Americas, otherwise known as the Clovis First theory.
Recent radiocarbon dating of Clovis sites has yielded ages of 11.1k to 10.7k 14C years BP (13k to 12.6k cal years BP), somewhat later than dates derived from older techniques.[70] The re-evaluation of earlier radiocarbon dates led to the conclusion that no fewer than 11 of the 22 Clovis sites with radiocarbon dates are "problematic" and should be disregarded, including the type site in Clovis, New Mexico. Numerical dating of Clovis sites has allowed comparison of Clovis dates with dates of other archaeosites throughout the Americas, and of the opening of the ice-free corridor. Both lead to significant challenges to the Clovis First theory. The Monte Verde site of Southern Chile has been dated at 14.8k cal years BP.[32] The Paisley Cave site in eastern Oregon yielded a 14C date of 12.4k years (14.5k cal years) BP, on a coprolite with human DNA and 14C dates of 11.3k-11k (13.2k-12.9k cal years) BP on horizons containing western stemmed points.[71] Artifact horizons with non-Clovis lithic assemblages and pre-Clovis ages occur in eastern North America, although the maximum ages tend to be poorly constrained.[31][38]
Geological findings on the timing of the ice-free corridor also challenge the notion that Clovis and pre-Clovis human occupation of the Americas was a result of migration through that route following the Last Glacial Maximum. Pre-LGM closing of the corridor may approach 30k cal years BP and estimates of ice retreat from the corridor are in the range of 12 to 13k cal years BP.[11][12][13] Viability of the corridor as a human migration route has been estimated at 11.5k cal years BP, later than the ages of the Clovis and pre-Clovis sites.[13] Dated Clovis archaeosites suggest a south-to-north spread of the Clovis culture.[11]
Pre-Last Glacial Maximum migration into the interior has been proposed to explain pre-Clovis ages for archaeosites in the Americas,[25][30] although pre-Clovis sites such as Meadowcroft Rock Shelter,[31][38] Monte Verde,[32] and Paisley Cave have not yielded confirmed pre-LGM ages.
The interior route is consistent with the spread of the Na Dene language group and Subhaplogroup X2a into the Americas after the earliest paleoamerican migration.[46]
Pacific coastal route
Pacific models propose that people first reached the Americas via water travel, following coastlines from northeast Asia into the Americas. Coastlines are unusually productive environments because they provide humans with access to a diverse array of plants and animals from both terrestrial and marine ecosystems. While not exclusive of land-based migrations, the Pacific 'coastal migration theory' helps explain how early colonists reached areas extremely distant from the Bering Strait region, including sites such as Monte Verde in southern Chile and Taima-Taima in western Venezuela. Two cultural components were discovered at Monte Verde near the Pacific coast of Chile. The youngest layer is radiocarbon dated at 12,500 radiocarbon years (~14,000 cal BP) and has produced the remains of several types of seaweeds collected from coastal habitats. The older and more controversial component may date back as far as 33,000 years, but few scholars currently accept this very early component.
As the chronology of deglaciation in the interior and coastal regions of North America became better understood, the coastal migration hypothesis was advanced by Knute Fladmark as an alternative to the ice-free corridor hypothesis.[72] Debate on coastal versus interior migration for initial settlement has centered on evidence for chronology of initial settlement of Beringia,[19][34] interior North America,[11] the Pacific coast of the Americas,[32] and timing of the opening of coastal versus interior migration routes indicated by geological evidence.[11][13] Complicating the debate has been the absence of archaeological data from the coastal and interior migration routes from the periods when the initial migration is proposed to have occurred. A recent variation of the coastal migration hypothesis is the marine migration hypothesis, which proposes that migrants with boats settled in coastal refugia during deglaciation of the coast.[22][23] The proposed use of boats adds a measure of flexibility to the chronology of coastal migration, as a continuous ice-free coast (16k-15k cal years BP) would no longer be required. A coastal east Asian source population is integral to the marine migration hypothesis.[22][23]
In 2014, the autosomal DNA of a toddler from Montana, dated at 10.7k 14C years (12.5-12.7 cal years) BP was sequenced.[73] The DNA was taken from a skeleton referred to as Anzick-1, found in close association with several Clovis artifacts. The analysis yielded identification of the mtDNA as belonging to Subhaplogroup D4h3a, a rare subclade of D4h3 occurring along the west coast of the Americas, as well as geneflow related to the Siberian Mal'ta population. The data indicate that Anzick-1 is from a population directly ancestral to present South American and Central American Native American populations. Anzick-1 is less closely related to present North American Native American populations. D4h3a has been identified as a clade associated with coastal migration.[46]
The problems associated with finding archaeological evidence for migration during a period of lowered sea level are well known.[22][74] Sites related to the first migration are usually submerged, so the location of such sites is obscured. Certain types of evidence dependent on organic material, such as radiocarbon dating, may be destroyed by submergence. Wave action can destroy site structures and scatter artifacts along a prograding shoreline. Additionally, Pacific coastal conditions tend to be unstable due to steep unstable terrain, earthquakes, tsunamis, and volcanoes. Strategies for finding earliest migration sites include identifying potential sites on submerged paleoshorelines, seeking sites in areas uplifted either by tectonics or isostatic rebound, and looking for riverine sites in areas that may have attracted coastal migrants.[22][74] Otherwise, coastal archaeology is dependent on secondary evidence related to lifestyles and technologies of maritime peoples from sites similar to those that would be associated with the original migration.
Other coastal models, dealing specifically with the peopling of the Pacific Northwest and California coasts, have been advocated by archaeologists Knut Fladmark, Roy Carlson, James Dixon, Jon Erlandson, Ruth Gruhn, and Daryl Fedje. In a 2007 article in the Journal of Island and Coastal Archaeology, Erlandson and his colleagues proposed a corollary to the coastal migration theory—the "kelp highway hypothesis"—arguing that productive kelp forests supporting similar suites of plants and animals would have existed near the end of the Pleistocene around much of the Pacific Rim from Japan to Beringia, the Pacific Northwest, and California, as well as the Andean Coast of South America. Once the coastlines of Alaska and British Columbia had deglaciated about 16,000 years ago, these kelp forest (along with estuarine, mangrove, and coral reef) habitats would have provided an ecologically similar migration corridor, entirely at sea level, and essentially unobstructed.
A 2016 DNA analysis of plants and animals suggest a coastal route was feasible.[75][76]
East Asians: Paleoindians of the coast
The boat-builders from Southeast Asia (Austronesian peoples) may have been one of the earliest groups to reach the shores of North America.[77][78][79] One theory suggests people in boats followed the coastline from the Kurile Islands to Alaska down the coasts of North and South America as far as Chile.[8] 62[13] 54, 57. The Haida nation on the Queen Charlotte Islands off the coast of British Columbia may have originated from these early Asian mariners between 25,000 and 12,000 years ago. Early watercraft migration would also explain the habitation of coastal sites in South America such as Pikimachay Cave in Peru by 20,000 years ago (disputed) and Monte Verde in Chile by 13,000 years ago [6 30; 8 383].
'There was boat use in Japan 20,000 years ago,' says Jon Erlandson, a University of Oregon anthropologist. 'The Kurile Islands (north of Japan) are like stepping stones to Beringia,' the then continuous land bridging the Bering Strait. Migrants, he said, could have then skirted the tidewater glaciers in Canada right on down the coast. [7 64]'
Problems with evaluating coastal migration models
The coastal migration models provide a different perspective on migration to the New World, but they are not without their own problems. One such problem is that global sea levels have risen over 120 metres (390 ft)[80] since the end of the last glacial period, and this has submerged the ancient coastlines that maritime people would have followed into the Americas. Finding sites associated with early coastal migrations is extremely difficult—and systematic excavation of any sites found in deeper waters is challenging and expensive. On the other hand, there is evidence of marine technologies found in the hills of the Channel Islands of California, circa 10,000 BCE.[81] If there was an early pre-Clovis coastal migration, there is always the possibility of a "failed colonization". Another problem that arises is the lack of hard evidence found for a "long chronology" theory. No sites have yet produced a consistent chronology older than about 12,500 radiocarbon years (~14,500 calendar years) , but research has been limited in South America related to the possibility of early coastal migrations.
Y-DNA among South American and Alaskan natives
The micro-satellite diversity and distribution of a Y lineage specific to South America suggest that certain Amerindian populations became isolated after the initial colonization of their regions.[82] The Na-Dené, Inuit and Indigenous Alaskan populations exhibit haplogroup Q (Y-DNA) mutations, but are distinct from other indigenous Amerindians with various mtDNA and autosomal DNA (atDNA) mutations.[83][84][85] This suggests that the earliest migrants into the northern extremes of North America and Greenland derived from later migrant populations.[86][87]
Other hypotheses
There exist a number of theories for pre-Columbian trans-oceanic migrations into the Americas.
See also
- Bluefish Caves
- Little John (archeological site)
- Dené–Yeniseian languages - proposed family of languages spoken by indigenous peoples of Asia and North America
- Early human migrations
- Historical migration
- History of Mesoamerica (Paleo-Indian)
- List of countries and islands by first human settlement
- Making North America (2015 PBS film)
- Norse colonization of the Americas
- Olmec alternative origin speculations
- Origins of Paleoindians
- Pendejo Cave
- Paleo-Indians period (Canada)
- Pre-Columbian trans-oceanic contact
- Solutrean hypothesis
- Recent African origin of modern humans
References
- ↑ Fagan, Brian M.; Durrani, Nadia (2016). World Prehistory: A Brief Introduction. Routledge. p. 124. ISBN 978-1-317-34244-1.
- 1 2 Goebel, Ted; Waters, Michael R.; O'Rourke, Dennis H. (2008). "The Late Pleistocene dispersal of modern humans in the Americas" (PDF). Science. 319 (5869): 1497–1502. doi:10.1126/science.1153569. PMID 18339930. Archived from the original on 2014-01-02. Retrieved 2010-02-05.
- ↑ Zimmer, Carl (January 3, 2018). "In the Bones of a Buried Child, Signs of a Massive Human Migration to the Americas". The New York Times. Retrieved January 3, 2018.
- ↑ Moreno-Mayer, J. Victor; et al. (2018). "Terminal Pleistocene Alaskan genome reveals first founding population of Native Americans". Nature. 553 (7687): 203–07. doi:10.1038/nature25173. Retrieved January 3, 2018.
- ↑ Nicole Waguespack (2012). "Early Paleoindians, from Colonization to Folsom". In Timothy R. Pauketat. The Oxford Handbook of North American Archaeology. Oxford University Press. pp. 86–95. ISBN 978-0-19-538011-8.
- ↑ Vicki Cummings; Peter Jordan; Marek Zvelebil (24 April 2014). The Oxford Handbook of the Archaeology and Anthropology of Hunter-Gatherers. OUP Oxford. p. 572. ISBN 978-0-19-102526-6.
- ↑ Ash, Patricia J.; Robinson, David J. (2011). The Emergence of Humans: An Exploration of the Evolutionary Timeline. John Wiley & Sons. p. 289. ISBN 978-1-119-96424-7.
- 1 2 Gremillion, David H.; Auton, Adam; Falush, Daniel (2008-09-25). "Archaeolog: Pre Siberian Human Migration to America: Possible validation by HTLV-1 mutation analysis". PLoS Genetics. Traumwerk.stanford.edu. 4 (5): e1000078. doi:10.1371/journal.pgen.1000078. PMC 2367454. PMID 18497854. Archived from the original on 2009-07-10. Retrieved 2010-10-12.
- ↑ Waters, M. R.; Forman, S. L.; Jennings, T. A.; Nordt, L. C.; Driese, S. G.; Feinberg, J. M.; Keene, J. L.; Halligan, J.; Lindquist, A.; Pierson, J.; Hallmark, C. T.; Collins, M. B.; Wiederhold, J. E. (24 March 2011). "The Buttermilk Creek Complex and the Origins of Clovis at the Debra L. Friedkin Site, Texas". Science. 331 (6024): 1599–1603. doi:10.1126/science.1201855.
- 1 2 3 4 5 6 7 8 Brigham-Grette, Julie, Lozhkin, Anatoly V., Anderson, Patricia M., and Glushkova, Olga Y., (2004-09-16). "Western Beringia before and during the Last Glacial Maximum (Chapter 2),". in Madsen, David B. (editor) Entering America: Northeast Asia and Beringia Before the Last Glacial Maximum, University of Utah Press, 2004, 486p. ISBN 0-87480-786-7. ISBN 9780874807868.
- 1 2 3 4 5 6 7 8 9 Pinsker, Lisa. "Geotimes - February 2004 - The Ice-Free Corridor Revisited".
- 1 2 3 4 5 Jackson L. E. Jr.; Phillips F. M.; Shimamura K.; Little E.C. (1997). "Cosmogenic 36Cl dating of the Foothills Erratics train, Alberta, Canada". Geology. 25 (3): 195–198. doi:10.1130/0091-7613(1997)025<0195:ccdotf>2.3.co;2.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 Mandryk, Carole A.S., Josenhans, Heiner, Fedje, Daryl W., and Mathewes, Rolf W. (2001). "Late Quaternary paleoenvironments of Northwestern North America: implications for inland versus coastal migration routes" (PDF). Quaternary Science Reviews. 20: 301–314. doi:10.1016/s0277-3791(00)00115-3.
- 1 2 Dyke, A S; Moore, A; Robertson, L., Deglaciation of North America, Geological Survey of Canada, Open File 1574, 2003
- 1 2 Booth, Derek B., Troost, Kathy Goetz, Clague, John J., and Waitt, Richard B. (2003). "The cordilleran ice sheet" (PDF). Developments in Quaternary Sciences. Developments in Quaternary Sciences. 1: 17–43. doi:10.1016/S1571-0866(03)01002-9. ISBN 9780444514707.
- 1 2 Blaise, B., Clague, J.J., and Mathewes, R.W., (1990). "Time of maximum Late Wisconsin glaciation, west coast of Canada". Quaternary Research. 34 (3): 282–295. doi:10.1016/0033-5894(90)90041-i.
- ↑ Misarti, Nicole, Finney, Bruce P., Jordan, James W., Maschner, Herbert D.G., Addison, Jason A., Shapley, Mark D., Krumhardt, Andrea, and Beget, James E., (2012). "Early retreat of the Alaska Peninsula Glacier Complex and the implications for coastal migrations of First Americans". Quaternary Science Reviews. 48: 1–6. doi:10.1016/j.quascirev.2012.05.014.
- 1 2 3 4 5 Clague John J., Mathewes, Rolf w., and Ager, Thomas A., (2004-09-16). "Western Beringia before and during the Last Glacial Maximum (Chapter 3),". Environments of Northwestern North America before the Last Glacial Maximum,in Madsen, David B. (editor) Entering America: Northeast Asia and Beringia Before the Last Glacial Maximum, University of Utah Press, 2004, 486p. ISBN 0-87480-786-7. ISBN 9780874807868.
- 1 2 3 4 5 6 7 Vasil'ev, Sergey A., Kuzmin, Yaroslav V., Orlova, Lyubov A., and Vyacheslav Dementiev, N. (2002). "Radiocarbon-based chronology of the Paleolithic in Siberia and its relevance to the peopling of the New World". Radiocarbon. 44 (2): 503–530. doi:10.1017/s0033822200031878.
- 1 2 3 Graf, Kelly E. (2009), "Modern human colonization of the mammoth steppe: a view from south-central Siberia" (PDF), Sourcebook of Paleolithic Transitions, Springer, in Camps, M, and Chauhan, P (eds.): 479–501, doi:10.1007/978-0-387-76487-0_32, ISBN 978-0-387-76478-8
- 1 2 3 4 5 Fedje, Daryl W., Mackie, Quentin, Dixon, E. James, and Heaton, Timothy H., (2004-09-16). Archaeological visibility along the northern northwest coast (Chapter 4),in Madsen, David B. (editor) Entering America: Northeast Asia and Beringia Before the Last Glacial Maximum, University of Utah Press, 2004, 486p. ISBN 0-87480-786-7. ISBN 9780874807868.
- 1 2 3 4 5 6 7 Erlandson, Jon M. & Braje, Todd J., (2011), "From Asia to the Americas by boat? Paleogeography, paleoecology, and stemmed points of the northwest Pacific" (PDF), Quaternary International, 239: 28–37, doi:10.1016/j.quaint.2011.02.030
- 1 2 3 4 Erlandson, Jon M., Graham, Michael H., Bourque, Bruce J., Corbett, Debra, Estes, James A., and Steneck, Robert S. (2007). "The Kelp highway hypothesis: marine ecology, the coastal migration theory, and the peopling of the Americas". The Journal of Island and Coastal Archaeology. 2 (2): 161–174. doi:10.1080/15564890701628612.
- ↑ Phillip M. White (2006). American Indian chronology: chronologies of the American mosaic. Greenwood Publishing Group. p. 1. ISBN 978-0-313-33820-5.
- 1 2 3 4 Wells, Spencer; Read, Mark (2002). The Journey of Man - A Genetic Odyssey (Digitised online by Google books). Random House. pp. 138–140. ISBN 0-8129-7146-9.
- ↑ Stefan Lovgren (March 13, 2008). "Americas Settled 15,000 Years Ago, Study Says".
- 1 2 3 4 Bonatto, Sandro L.; Salzano, Francisco M. (1997). "A single and early migration for the peopling of the Americas supported by mitochondrial DNA sequence data". Proceedings of the National Academy of Sciences. 94 (5): 1866–1871. doi:10.1073/pnas.94.5.1866. PMC 20009. PMID 9050871.
- 1 2 Cinq-Mars J. (1979). "Bluefish Cave 1: A Late Pleistocene Eastern Beringian Cave Deposit in the Northern Yukon,". Canadian Journal of Archaeology (3): 1–32.
- 1 2 Bonnichsen, Robson, Critical arguments for Pleistocene artifacts from the Old Crow basin, Yukon: a preliminary statement (p.102-118), in Bryan, Alan L., editor Early Man in America from a Circum-Pacific Perspective. Edmonton: Archaeological Researches International (Occasional Papers No. 1), Department of Anthropology, University of Alberta, 1978
- 1 2 3 "Journey of mankind". Bradshaw Foundation.
- 1 2 3 4 Goodyear, Albert C., Evidence of Pre-Clovis sites in the eastern United States, in Paleoamerican Origins: Beyond Clovis, 2005, Texas A&M University, Center for the Study of the First Americans, p. 103-112
- 1 2 3 4 5 Dillehay, Thomas (2000). The Settlement of the Americas: A New Prehistory. New York: Basic Books. ISBN 0-465-07669-6.
- ↑ Figure 4 of Kitchen Andrew (2008). "A Three-Stage Colonization Model for the Peopling of the Americas". PLoS ONE. 3 (2): e1596. doi:10.1371/journal.pone.0001596.
- 1 2 3 Goebel, Ted & Buvit, Ian. (2011-04-01), From the Yenisei to the Yukon: Interpreting Lithic Assemblages in Late Pleistocene/Early Holocene Beringia, Center for the Study of the First Americans, 2011, ISBN 9781603443845
- ↑ G.M. Santos, M.I. Bird, F. Parenti, L.K Fifield, N. Guidon, P.A Hausladen, "A revised chronology of the lowest occupation layer of Pedra Furada Rock Shelter, Piauı́, Brazil: the Pleistocene peopling of the Americas", Quaternary Science Reviews Volume 22, Issues 21–22, November–December 2003, pp. 2303-2310, doi:0.1016/S0277-3791(03)00205-1.
- ↑ van Vark GN, Kuizenga D, Williams FL (June 2003). "Kennewick and Luzia: lessons from the European Upper Paleolithic". American Journal of Physical Anthropology. 121 (2): 181–4, discussion 185–8. doi:10.1002/ajpa.10176. PMID 12740961. Fiedel, Stuart J. (2004). "The Kennewick Follies: 'New' Theories about the Peopling of the Americas". Journal of Anthropological Research. 60 (1): 75–110. JSTOR 3631009. González-José R, Bortolini MC, Santos FR, Bonatto SL (October 2008). "The peopling of America: craniofacial shape variation on a continental scale and its interpretation from an interdisciplinary view". American Journal of Physical Anthropology. 137 (2): 175–87. doi:10.1002/ajpa.20854. PMID 18481303.
- ↑ Gibbon, Guy E; Ames, Kenneth M (1998). Archaeology of Prehistoric Native America: An Encyclopedia|work=. By Guy E. Gibbon, Kenneth M. Ames (1998) ISBN 978-0-8153-0725-9.
- 1 2 3 Adovasio, J. M, J. Donahue, and R. Stuckenrath (1990). "The Meadowcroft Rockshelter Radiocarbon Chronology 1975–1990". American Antiquity. 55 (2): 348–54. doi:10.2307/281652. JSTOR 281652.
- ↑ Holen, Steven R.; Deméré, Thomas A.; Fisher, Daniel C.; Fullagar, Richard; Paces, James B.; Jefferson, George T.; Beeton, Jared M.; Cerutti, Richard A.; Rountrey, Adam N.; Vescera, Lawrence; Holen, Kathleen A. (2017). "A 130,000-year-old archaeological site in southern California, USA". Nature. 544 (7651): 479–483. doi:10.1038/nature22065. PMID 28447646. Retrieved 30 April 2017.
- ↑ Rincon, Paul (26 April 2017). "First Americans claim sparks controversy". BBC News. Retrieved 30 April 2017. Michael R. Waters commented that "To demonstrate such early occupation of the Americas requires the presence of unequivocal stone artefacts. There are no unequivocal stone tools associated with the bones... this site is likely just an interesting paleontological locality." Chris Stringer said that "extraordinary claims require extraordinary evidence - each aspect requires the strongest scrutiny," adding that "High and concentrated forces must have been required to smash the thickest mastodon bones, and the low energy depositional environment seemingly provides no obvious alternative to humans using the heavy cobbles found with the bones.
- ↑ Pitulko, V. V.; Nikolsky, P. A.; Girya, E. Yu; Basilyan, A. E.; Tumskoy, V. E.; Koulakov, S. A.; Astakhov, S. N.; Pavlova, E. Yu; Anisimov, M. A. (2 January 2004). "The Yana RHS Site: Humans in the Arctic Before the Last Glacial Maximum". Science. 303 (5654): 52–56. doi:10.1126/science.1085219. ISSN 0036-8075. PMID 14704419. Retrieved 30 April 2017.
- 1 2 3 4 5 6 7 Tamm E, Kivisild T, Reidla M, Metspalu M, Smith DG; et al. (2007). "Beringian Standstill and Spread of Native American Founders". PLoS ONE. 2 (9): e829. doi:10.1371/journal.pone.0000829. PMC 1952074. PMID 17786201.
- 1 2 3 4 Kitchen, Andrew, Miyamoto, Michal M., and Mulligan, Connie J., (2008). "A Three-Stage Colonization Model for the Peopling of the Americas". PLoS ONE. 3 (2): e1596. doi:10.1371/journal.pone.0001596. PMC 2223069. PMID 18270583.
- ↑ P. Skoglund, D. Reich, "A genomic view of the peopling of the Americas", Curr Opin Genet Dev. 2016 Dec; 41: 27–35, doi: 10.1016/j.gde.2016.06.016. "Recently, we carried out a stringent test of the null hypothesis of a single founding population of Central and South Americans using genome-wide data from diverse Native Americans. We detected a statistically clear signal linking Native Americans in the Amazonian region of Brazil to present-day Australo-Melanesians and Andaman Islanders (‘Australasians’). Specifically, we found that Australasians share significantly more genetic variants with some Amazonian populations—including ones speaking Tupi languages—than they do with other Native Americans. We called this putative ancient Native American lineage “Population Y” after Ypykuéra, which means ‘ancestor’ in the Tupi language family."
- 1 2 3 Brian M. Kemp, Ripan S. Malhi, John McDonough, Deborah A. Bolnick, Jason A. Eshleman, Olga Rickards, Cristina Martinez-Labarga, John R. Johnson, Joseph G. Lorenz, E. James Dixon, Terence E. Fifield, Timothy H. Heaton, Rosita Worl, and David Glenn Smith (2007). "Genetic Analysis of Early Holocene Skeletal Remains From Alaska and its Implications for the Settlement of the Americas" (PDF). Am J Phys Anthropol. 132 (4): 605–21. doi:10.1002/ajpa.20543. PMID 17243155.
- 1 2 3 4 5 6 Perego, Ugo A., Achilli, Alessandro, Angerhofer, Norman, Accetturo, Matteo, Pala, Maria, Olivieri, Anna, Kashani, Baharak H., Ritchie, Kathleen H., Scozzari, Rosaria, Kong, Qing-Peng, Myres, Natalie M., Salas, Antonio, Semino, Ornella, Bandelt, Hans-Jürgen, Woodward, Scott R., and Torroni, Antonio (2009). "Distinctive Paleo-Indian Migration Routes from Beringia Marked by Two Rare mtDNA Haplogroups". Current Biology. 19 (1): 1–8. doi:10.1016/j.cub.2008.11.058. PMID 19135370.
- 1 2 Derenko M, Malyarchuk B, Grzybowski T, Denisova G, Rogalla U, et al. (2010). "Origin and Post-Glacial Dispersal of Mitochondrial DNA Haplogroups C and D in Northern Asia". PLoS ONE. 5 (12): e15214. doi:10.1371/journal.pone.0015214. PMC 3006427. PMID 21203537.
- ↑ Maria-Catira Bortolini; Francisco M. Salzano; Mark G. Thomas; Steven Stuart; Selja P. K. Nasanen; Claiton H. D. Bau; Mara H. Hutz; Zulay Layrisse; Maria L. Petzl-Erler; Luiza T. Tsuneto; Kim Hill; Ana M. Hurtado; Dinorah Castro-de-Guerra; Maria M. Torres; Helena Groot; Roman Michalski; Pagbajabyn Nymadawa; Gabriel Bedoya; Neil Bradman; Damian Labuda; Andres Ruiz-Linares. (2003). "Y-chromosome evidence for differing ancient demographic histories in the Americas" (PDF). American Journal of Human Genetics. 73 (3): 524–539. doi:10.1086/377588. PMC 1180678. PMID 12900798.
- 1 2 3 Schurr, Theodore G. "Mitochondrial DNA and the Peopling of the New World" (PDF). American Scientist Online May–June 2000. Retrieved May 13, 2015.
- 1 2 Zakharov, I. A., Derenko, M. V., Maliarchuk, B. A., Dambueva I. K., Dorzhu, C. M., and Rychkov, S. Y. (April 2004). "Mitochondrial DNA variation in the aboriginal populations of the Altai-Baikal region: implications for the genetic history of North Asia and America". Ann. N. Y. Acad. Sci. 1011: 21–35. doi:10.1196/annals.1293.003. PMID 15126280.
- 1 2 3 4 5 Starikovskaya, Elena B., Sukernik, Rem I., Derbeneva, Olga A., Volodko, Natalia A., Ruiz-Pesini, Eduardo, Torroni, Antonio, Brown, Michael D., Lott, Marie T., Hosseini, Seyed H., Huoponen, Kirsi, and Wallace, Douglas C. (January 2005). "Mitochondrial DNA diversity in indigenous populations of the southern extent of Siberia, and the origins of Native American haplogroups". Ann. Hum. Genet. 69 (Pt 1): 67–89. doi:10.1046/j.1529-8817.2003.00127.x. PMC 3905771. PMID 15638829.
- 1 2 3 4 Adachi N, Shinoda K, Umetsu K, et al. (2009). "Mitochondrial DNA analysis of Jomon skeletons from the Funadomari site, Hokkaido, and its implication for the origins of Native American". Am J Phys Anthropol. 138 (3): 255–65. doi:10.1002/ajpa.20923. PMID 18951391.
- ↑ Adachi, N., Shinoda, K. i., Umetsu, K., Kitano, T., Matsumura, H., Fujiyama, R., Sawada, J. and Tanaka, M. (2011). "Mitochondrial DNA analysis of Hokkaido Jomon skeletons: Remnants of archaic maternal lineages at the southwestern edge of former Beringia". American Journal of Physical Anthropology. 146 (3): 346–360. doi:10.1002/ajpa.21561. PMID 21953438.
- ↑ Li, Hong-Chuan, Robert J. Biggar, Wendell J. Miley, Elizabeth M. Maloney, Beverley Cranston, Barrie Hanchard and Michie Hisada (2004). "Provirus load in breast milk and risk of mother-to-child transmission of Human T Lymphotropic Virus Type I". The Journal of Infectious Diseases. 190 (7): 1275–1278. doi:10.1086/423941.
- 1 2 3 4 Verdonck K, González E, Van Dooren S, Vandamme AM, Vanham G, and Gotuzzo E. (Apr 2007). "Human T-lymphotropic virus 1: recent knowledge about an ancient infection". Lancet Infect Dis. 7 (4): 266–81. doi:10.1016/S1473-3099(07)70081-6. PMID 17376384.
- ↑ Gessain, A., Gallo, R. C., and Franchini, G. (Apr 1992). "Low degree of human T-cell leukemia/lymphoma virus type I genetic drift in vivo as a means of monitoring viral transmission and movement of ancient human populations". J Virol. 66 (4): 2288–95. PMC 289023. PMID 1548762.
- ↑ Ishida T, Yamamoto K, Omoto K, Iwanaga M, Osato T, and Hinuma Y. (Sep 1985). "Prevalence of a human retrovirus in native Japanese: evidence for a possible ancient origin". J Infect. 11 (2): 153–7. doi:10.1016/s0163-4453(85)92099-7. PMID 2997332.
- 1 2 Miura, t., T Fukunaga, T Igarashi, M Yamashita, E Ido, S Funahashi, T Ishida, K Washio, S Ueda, K Hashimoto (Feb 1994). "Phylogenetic subtypes of human T-lymphotropic virus type I and their relations to the anthropological background". Proc Natl Acad Sci U S A. 91 (3): 1124–7. doi:10.1073/pnas.91.3.1124. PMC 521466. PMID 8302841.
- ↑ Picard, F. J., M. B. Coulthart, J. Oger, E. E. King, S. Kim, J. Arp, G. P. Rice, and G. A. Dekaban (Nov 1995). "Human T-lymphotropic virus type 1 in coastal natives of British Columbia: phylogenetic affinities and possible origins". J Virol. 69 (11): 7248–56. PMC 189647. PMID 7474147.
- ↑ Li, H.C., Fujiyoshi T., Lou, H., Yashiki, S., Sonoda, S., Cartier, L., Nunez, L., Munoz, I., Horai, S., and Tajima, K. (Dec 1999). "The presence of ancient human T-cell lymphotropic virus type I provirus DNA in an Andean mummy". Nat Med. 5 (12): 1428–32. doi:10.1038/71006. PMID 10581088.
- 1 2 Coulthart, Michael B., David Posada, Keith A. Crandall, and Gregory A. Dekaband (Mar 2006). "On the phylogenetic placement of human T cell leukemia virus type 1 sequences associated with an Andean mummy". Infect Genet Evol. 6 (2): 91–6. doi:10.1016/j.meegid.2005.02.001. PMC 1983367. PMID 16503510.
- 1 2 Gonzaleza, Silvia; David Huddart, Isabel Israde -Alcántara, Gabriela Domínguez-Vázquez, James Bischoff, Nicholas Felstead (2015). "Paleoindian sites from the Basin of Mexico: Evidence from stratigraphy, tephrochronology and dating". Quaternary International. 363: 4–19. doi:10.1016/j.quaint.2014.03.015.
- 1 2 González-José, Rolando; Antonio González-Martín, Miquel Hernández, Héctor M. Pucciarelli, Marina Sardi, Alfonso Rosales, and Silvina Van der Molen (4 September 2003). "Craniometric evidence for Palaeoamerican survival in Baja California". Nature. 425 (6953): 62–65. doi:10.1038/nature01816. PMID 12955139.
- ↑ Dillehay, Thomas D. (4 September 2003). "Tracking the first Americans" (PDF). Nature. 425 (6953): 23–24. doi:10.1038/425023a.
- ↑ Fiedel, Stuart J. (Spring 2004). "The Kennewick follies: "new" theories about the peopling of the Americas". Journal of Anthropological Research. 60 (1). JSTOR 3631009.
- ↑ Chatters, James C., Douglas J. Kennett, Yemane Asmerom, Brian M. Kemp, Victor Polyak, Alberto Nava Blank, Patricia A. Beddows, Eduard Reinhardt, Joaquin Arroyo-Cabrales, Deborah A. Bolnick, Ripan S. Malhi, Brendan J. Culleton, Pilar Luna Erreguerena, Dominique Rissolo, Shanti Morell-Hart, and Thomas W. Stafford Jr. (2014). "Late Pleistocene Human Skeleton and mtDNA Link Paleoamericans and Modern Native Americans" (PDF). Science. 344 (6185): 750–754. doi:10.1126/science.1252619. Archived from the original (PDF) on 2015-07-13.
- ↑ de Azvedo, Soledad; Bortolini, Maria C.; Bonatto, Sandro L.; Hunemeier, Tabita; Santos, Fabrıcio R. S; Gonzalez-Jose, Rolando (January 2015). "Ancient Remains and the First Peopling of the Americas: Reassessing the Hoyo Negro Skull". American Journal of Physical Anthropology. 148 (3): 514–521. doi:10.1002/ajpa.22801.
- ↑ de Azevedo S.; et al. (2015). ""The first human settlement of the New World " A closer look at craniofacial variation and evolution of early and late Holocene Native American groups" (PDF). Quaternary International. 431: 152–167. doi:10.1016/j.quaint.2015.11.012.
- ↑ Seong, C. (2008). "Tanged points, microblades and late paleolithic hunting in Korea". Antiquity. 82 (318): 871–883. doi:10.1017/s0003598x00097647.
- ↑ Waters, M. R. & Stafford, T. W. (2007). "Redefining the age of Clovis: implications for the peopling of the Americas". Science. 315 (5815): 1122–1126. doi:10.1126/science.1137166. PMID 17322060.
- ↑ Jenkins, Dennis L., Loren G. Davis, Thomas W. Stafford Jr., Paula F. Campos, Bryan Hockett, George T. Jones, Linda Scott Cummings, Chad Yost, Thomas J. Connolly, Robert M. Yohe II, Summer C. Gibbons, Maanasa Raghavan, Morten Rasmussen, Johanna L. A. Paijmans, Michael Hofreiter, Brian M. Kemp, Jodi Lynn Barta, Cara Monroe, M. Thomas, P. Gilbert, and Eske Willerslev (13 July 2012). "Clovis Age Western Stemmed Projectile Points and Human Coprolites at the Paisley Caves" (PDF). Science. 337 (6091): 223–228. doi:10.1126/science.1218443. PMID 22798611.
- ↑ Fladmark, K.R. (1979). "Routes: alternate migration corridors for early man in North America". American Antiquity. 44: 55–69. doi:10.2307/279189. JSTOR 279189.
- ↑ Rasmussen, Morten, Sarah L. Anzick, Michael R. Waters, Pontus Skoglund, Michael DeGiorgio, Thomas W. Stafford Jr.; et al. (February 2014). "The genome of a Late Pleistocene human from aClovis burial site in western Montana". Nature. 506 (7487): 225–229. doi:10.1038/nature13025. PMC 4878442. PMID 24522598.
- 1 2 Pinsker, Lisa. "Geotimes - February 2004 - Quest for the Lost Land".
- ↑ "Plant and animal DNA suggests first Americans took the coastal route". Nature. doi:10.1038/536138a. Retrieved 11 August 2016.
- ↑ Humans may have taken different path into Americas than thought Arctic passage wouldn’t have provided enough food for the earliest Americans’ journey by Thomas Summer, published in "Science News" on August 10, 2016
- ↑ Gunn, Bee; Luc Baudouin; Kenneth M. Olsen (2011). "Independent Origins of Cultivated Coconut (Cocos nucifera L.) in the Old World Tropics". PLoS ONE. 6 (6): e21143. doi:10.1371/journal.pone.0021143. PMC 3120816. PMID 21731660.
- ↑ Van Tilburg, Jo Anne. 1994. Easter Island: Archaeology, Ecology and Culture. Washington D.C.: Smithsonian Institution Press
- ↑ Langdon, Robert. The Bamboo Raft as a Key to the Introduction of the Sweet Potato in Prehistoric Polynesia, The Journal of Pacific History, Vol. 36, No. 1, 2001
- ↑ Gornitz, Vivian (January 2007). "Sea Level Rise, After the Ice Melted and Today". Goddard Institute for Space Studies. NASA. Retrieved 23 April 2015.
- ↑ "California islands give up evidence of early seafaring: Numerous artifacts found at late Pleistocene sites on the Channel Islands," ScienceDaily, March 4, 2011. https://www.sciencedaily.com/releases/2011/03/110303141540.htm
- ↑ "Summary of knowledge on the subclades of Haplogroup Q". Genebase Systems. 2009. Archived from the original on 2011-05-10. Retrieved 2009-11-22.
- ↑ Ruhlen M (November 1998). "The origin of the Na-Dene". Proceedings of the National Academy of Sciences of the United States of America. 95 (23): 13994–6. doi:10.1073/pnas.95.23.13994. PMC 25007. PMID 9811914.
- ↑ Zegura SL, Karafet TM, Zhivotovsky LA, Hammer MF; Karafet; Zhivotovsky; Hammer (January 2004). "High-resolution SNPs and microsatellite haplotypes point to a single, recent entry of Native American Y chromosomes into the Americas". Molecular Biology and Evolution. 21 (1): 164–75. doi:10.1093/molbev/msh009. PMID 14595095.
- ↑ Saillard, Juliette Saillard, Peter Forster, Niels Lynnerup1, Hans-Jürgen Bandelt and Søren Nørby; Forster, Peter; Lynnerup, Niels; Bandelt, Hans-Jürgen; Nørby, Søren (2000). "mtDNA Variation among Greenland Eskimos. The Edge of the Beringian Expansion". The American Journal of Human Genetics. 67 (3): 718–726. doi:10.1086/303038. PMC 1287530. PMID 10924403. Retrieved 2009-11-22.
- ↑ Schurr, Theodore G. (2004). "The peopling of the New World - Perspectives from Molecular Anthropology". Annual Review of Anthropology. 33: 551&ndash, 583. doi:10.1146/annurev.anthro.33.070203.143932.
- ↑ Torroni, Antonio; Schurr, Theodore G.; Yang, Chi-Chuan; Szathmary, Emöke J. E.; Williams, Robert C.; et al. "Native American Mitochondrial DNA Analysis Indicates That the Amerind and the Nadene Populations Were Founded by Two Independent Migrations". Genetics Society of America. 130: 153–162. Retrieved 2009-11-28.
{kind=link}
{kind=link}
Bibliography
- Adovasio, J. M., with Jake Page. The First Americans: In Pursuit of Archaeology's Greatest Mystery. New York: Random House, 2002.
- Bradley, B.; Stanford, D. (2004). "The North Atlantic ice-edge corridor: a possible Palaeolithic route to the New World". World Archaeology. 36 (4): 459–478. doi:10.1080/0043824042000303656.
- Bradley, B.; Stanford, D. (2006). "The Solutrean-Clovis connection: reply to Straus, Meltzer and Goebel". World Archaeology. 38 (4): 704–714. doi:10.1080/00438240601022001. JSTOR 40024066.
- Dennis J. Stanford, Bruce Bradely, Pre-Clovis First Americans: The Origin of America's Clovis Culture (University of California Press, 2012). ISBN 978-0-520-22783-5
- Dennis J. Stanford, Bruce A. Bradley, Across Atlantic Ice: The Origin of America's Clovis Culture (University of California Press (2012). ISBN 978-0-520-22783-5
- Diamond, Jared, Guns, germs and steel. A short history of everybody for the last 13,000 years, 1997.
- Dixon, E. James. Quest for the Origins of the First Americans. University of New Mexico Press. 1993.
- Dixon, E. James. Bones, Boats, and Bison: the Early Archeology of Western North America. University of New Mexico Press. 1993, 1999??
- Erlandson, Jon M. Early Hunter-Gatherers of the California Coast. Plenum Press. 1994.
- Erlandson, Jon M (2001). "The Archaeology of Aquatic Adaptations: Paradigms for a New Millennium". Journal of Archaeological Research. 9 (4): 287–350. doi:10.1023/a:1013062712695.
- Erlandson, Jon M. Anatomically Modern Humans, Maritime Migrations, and the Peopling of the New World. In The First Americans: The Pleistocene Colonization of the New World, edited by N. Jablonski, 2002. pp. 59–92. Memoirs of the California Academy of Sciences. San Francisco.
- Erlandson, Jon. M.; Graham, M. H.; Bourque, Bruce J.; Corbett, Debra; Estes, James A.; Steneck, R. S. (2007). "The Kelp Highway Hypothesis: Marine Ecology, The Coastal Migration Theory, and the Peopling of the Americas". Journal of Island and Coastal Archaeology. 2 (2): 161–174. doi:10.1080/15564890701628612.
- Eshleman, Jason A.; Malhi, Ripan S.; Glenn Smith, David (2003). "Mitochondrial DNA Studies of Native Americans: Conceptions and Misconceptions of the Population Prehistory of the Americas". Evolutionary Anthropology. 12: 7–18. doi:10.1002/evan.10048.
- Fedje; Christensen (1999). "Modeling Paleoshorelines and Locating Early Holocene Coastal Sites in Haida Gwaii". American Antiquity. 64 (4): 635–652. doi:10.2307/2694209.
- Greenman, E. F. (1963). "The Upper Palaeolithic and the New World". Current Anthropology. 4: 41–66. doi:10.1086/200337.
- Hey, Jody (2005). "On the Number of New World Founders: A Population Genetic Portrait of the Peopling of the Americas". PLOS Biology. 3 (6): e193. doi:10.1371/journal.pbio.0030193. PMC 1131883. PMID 15898833.
- Nina G. Jablonski (2002). The First Americans: The Pleistocene Colonization of the New World. California Academy of Sciences. ISBN 978-0-940228-50-4.
- Jones, Peter N. Respect for the Ancestors: American Indian Cultural Affiliation in the American West. Boulder, Colorado: Bauu Press. 2004, 2005.
- Lauber, Patricia. Who Came First? New Clues to Prehistoric Americans. Washington, D.C.: National Geographic Society, 2003.
- Matson and Coupland. The Prehistory of the Northwest Coast. Academic Press. New York. 1995.
- David J. Meltzer (2009). First Peoples in a New World: Colonizing Ice Age America. University of California Press. ISBN 978-0-520-94315-5.
- Snow, Dean R. "The First Americans and the Differentiation of Hunter-Gatherer Cultures." In Bruce G. Trigger and Wilcomb *E. Washburn, eds., The Cambridge History of the Native Peoples of the Americas, Volume I: North America (Cambridge: Cambridge University Press, 1996), 125-199.
- Spencer Wells (2002). The Journey of Man: A Genetic Odyssey. Princeton University Press. ISBN 0-691-11532-X.
- Evidence Supports Earlier Date for People in North America, April 4, 2008
External links
| Wikimedia Commons has media related to Indigenous nations of the Americas. |
- "The first Americans: How and when were the Americas populated?", Earth, January 2016
- "When Did Humans Come to the Americas?" - Smithsonian Magazine February 2013
- Ordering information and news items for The Dene–Yeniseian Connection; the 2011 2nd printing has corriagenda for 14 articles in the 2010 ist printing
- Journey of Man: A Genetic Odyssey (movie) on YouTube - by Spencer Wells - PBS and National Geographic Channel, 2003 - 120 Minutes, UPC/EAN: 841887001267
- Atlas of the Human Journey, Genographic Project, National Geographic
- Journey of Mankind - Genetic Map – Bradshaw Foundation
- The Paleoindian Period – United States Department of the Interior, National Park Service
- Alabama Archaeology: Prehistoric Alabama – The University of Alabama, Department of Archaeology
- The Paleoindian Database – The University of Tennessee, Department of Anthropology.
- Paleoindians and the Great Pleistocene Die-Off – American Academy of Arts and Sciences, National Humanities Center
| History |  | |
|---|---|---|
| Settlement | ||
| Societies | ||
| Related |
| |
| Lists |
| |
| Chronology |
| |
| ||
| Pre-history | .svg.png) | ||||
|---|---|---|---|---|---|
| Mythology/Religion | |||||
| European colonization | |||||
| Modern groups by country |
| ||||
| Related topics | |||||
| |||||