Haplogroup O-M176
Haplogroup O-M176
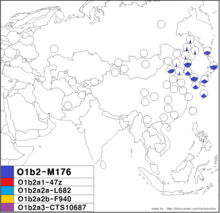 O1b2-M176 | |
|---|---|
| Possible time of origin | 28,400 [95% CI 26,000 <-> 30,800] years before present[1] |
| Coalescence age | 12,100 [95% CI 9,800 <-> 14,700] years before present (YFull[1][2]) |
| Possible place of origin | Korean Peninsula, Manchuria or a nearby part of northern East Asia[3][4] |
| Ancestor | O-P31 |
| Defining mutations | M176/SRY465, P49, 022454 |
| Highest frequencies |
Koreans, Japanese, Ryukyuans, Manchus
|
In human population genetics, Y-Chromosome haplogroups define the major lineages of direct paternal (male) lines back to a shared common ancestor in Africa. Haplogroup O-M176 (aka O-SRY465) is a human Y-chromosome DNA haplogroup. It is best known for its part in the settlement of Korea and Japan. It is a descendant of Haplogroup O-P31, and it has been estimated to share a most recent common ancestor with its nearest outgroup, Haplogroup O-K18, approximately 28,400 [95% CI 26,000 <-> 30,800] years before present.[1]
Distribution
Haplogroup O-M176 is found mainly in the northernmost parts of East Asia, from the Uriankhai and Zakhchin peoples of western Mongolia (Katoh 2004) to the Japanese of Japan, though it also has been detected sporadically in the Buryats (Jin 2003). It's been detected with moderate frequencies in Udegeys (Jin 2010) of southern Siberia, rarely among populations of Southeast Asia including Indonesia (Hammer 2006 and Jin 2003), the Philippines (Jin 2003), Thailand (Jin 2003), and Vietnam (Hammer 2006 and Jin 2003), and Micronesians (Hammer 2006). This haplogroup is found with its highest frequency and diversity values among modern populations of Japan and Korea and is rare in most populations in China. Among Han Chinese, it has been detected in some samples of Han Chinese from Beijing (1/51, Jin 2003 and Kim 2011)[5], Xi'an (1/34, Kim 2011)[5], Han Chinese in Taiwan (2/352 = 0.57%, including one of 34 Hakka people and one of 258 miscellaneous Han volunteers),[6] Han Chinese from East China sampled from the infertility clinic at the Affiliated Hospitals of Nanjing Medical University at Jiangsu (6/1147 = 0.52%, Lu 2009), Wuhan (1/160)[7], and South China outside of Jiangsu, Anhui, Zhejiang, and Shanghai (1/65).[8] Among ethnic minorities in China, haplogroup O-M176 has been detected with high frequency in samples of Koreans in China (Xue 2006 and Katoh 2004) and with generally much lower frequency among Manchus (Xue 2006, Katoh 2004, and Karafet 2001), Hezhe people,[9] Daurs,[9] Evenks,[10] Sibes (Xue 2006), Kham Tibetans,[11] and Hui.[12]
Subclade distribution
Paragroup O-M176*
Y-DNA that belongs to O-M176(xK10, F3356) has been found in an individual from Hiroshima,[2] an individual from Fukushima,[2] an individual from Beijing,[2] and 1% (7/706) of a sample of males collected in Seoul and Daejeon.[13]
O-M176(x47z) has been found in approximately 3.5%[14] to 9.9%[15] of Japanese males. However, most of those individuals probably belong to subclades of O-K10(x47z).
O-K10
The majority of extant members of O-M176 belong to the subclade O-K10 (or O-F3356). O-K10 subsumes the prolific subclades O-47z, which occurs with especially high frequency in Japan, and O-L682, which occurs with especially high frequency in Korea, in addition to a relatively rare subclade, O-K3, which has been found among Han Chinese in Hunan[2] and Manchus. O-L682 and O-K3 are linked by 18 SNPs that define the O-K4 clade, and thus their members are more closely related to one another by paternal lineage than any of them is related to any member of O-47z.
An example of Y-DNA that belongs to O-K10 but neither O-47z nor O-K4 has been found in an individual in Tokyo, Japan. This individual's Y-DNA is estimated to share a most recent common ancestor with O-47z and O-K4 roughly around the same time as the most recent common ancestor of those latter two clades, about 8,200 [95% CI 6,300 <-> 10,200] ybp.[1]
O-F3356(x47z, L682) has been found in 2% (14/706) of a sample of Koreans collected in Seoul and Daejeon, South Korea.[13] However, the status of these individuals' Y-DNA in regard to K4, K3, and phylogenetically equivalent SNPs has not been published.
O-47z
Haplogroup O-47z
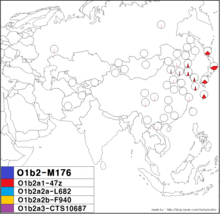 O1b2a1-47z | |
|---|---|
| Possible time of origin |
7,870 [95% CI 5,720–12,630] years ago (Hammer 2006) 8,200 [95% CI 6,300 <-> 10,200] ybp[1] |
| Possible place of origin | Japanese Archipelago(Hammer 2006) or Korean Peninsula(see Jin 2003) |
| Ancestor | O-M176 |
| Defining mutations | 47z |
| Highest frequencies |
Japanese, Ryukyuans, Koreans, Manchus
|
O-47z or O-CTS11986 is a subclade of O-K10. It is found with high frequency among the Japanese and Ryukyuan populations of Japan, and with lower frequency among Koreans.
Haplogroup O-47z has been detected in approximately 24% of males who speak a Japonic language, while it has not been found at all among Ainu males whose Y-DNA has been tested in two genetic studies (Tajima 2004, n=16; Hammer 2006, n=4). Based on the STR haplotype diversity within Haplogroup O-47z, it has been estimated in a study published in 2006 that this haplogroup has expanded from a single founder who has lived approximately 3,810 (95% CI 1,640 <–> 7,960) years before present in a model according to which continuous, pure exponential population growth is assumed.[10] In a paper published in 2016, the time to most recent common ancestor of a set of fifteen members of the O-47z clade, all from the JPT (Japanese in Tokyo, Japan) sample of the 1000 Genomes Project, was estimated to be 4,500 years using a relatively slow mutation rate (μ = 0.76 x 10–9 per bp per year as according to Qiaomei Fu et al. 2014) or 3,900 years using a relatively fast mutation rate (μ = 0.888 x 10–9 per bp per year as according to A. Helgason et al. 2015).[14] Haplogroup O-47z also has been found among samples of modern Koreans, though with low frequency in comparison to both the frequency of O-47z in samples of Japanese and the frequency of O-M176(x47z) in samples of Koreans.
O-K4
O-K4 is a subclade of O-K10. It includes at least two subclades, O-L682 and O-K3, which have been estimated to share a most recent common ancestor approximately 6,300 [95% CI 4,700 <-> 8,000] years before present.[2]
O-K3
Haplogroup O-F940
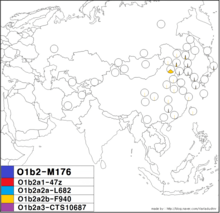 O1b2a2b-F940 | |
|---|---|
| Possible time of origin | 3,500 [2,000-13,000] |
| Possible place of origin | Manchuria or Korean Peninsula |
| Ancestor | O-F2868 |
| Defining mutations | CTS12145, F1912, F2206, F2703, F940, K3 |
| Highest frequencies |
Manchurian, Manchu Mongolian, Manchu Han
|
The O-K3 (or O-F940) lineage is a subclade of O-K4 that has been observed to date in two Han Chinese individuals from Hunan.[2]
O-L682
Haplogroup O-L682
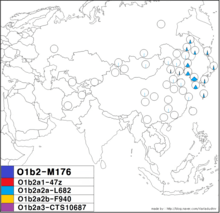 O1b2a2a-L682 | |
|---|---|
| Possible time of origin | 6,300 [95% CI 4,700 <-> 8,000] ybp[1] |
| Possible place of origin | Korean Peninsula or Manchuria |
| Ancestor | O-M176, O-F3356 |
| Defining mutations | L682 |
| Highest frequencies |
Koreans, Hezhen, Udege, Japanese, Ryukyuans, Manchus
|
The O-L682 subclade of O-K4 is believed to be related to Native Korean population. One study has found O-L682 Y-DNA in 19% (134/706) of Koreans sampled in Seoul and Daejeon.[13] O-L682 also has been found in Japanese in Tokyo, Okayama, Kōchi, and the USA, Chinese in Shanxi, Shandong, and Beijing,[2] and Nanai people in China. Its descendants appear to have begun rapidly increasing in number at approximately the same time as those of its distant cousin O-47z, perhaps 4,000 years ago.[2]
Phylogenetics
Phylogenetic history
Prior to 2002, there were in academic literature at least seven naming systems for the Y-Chromosome phylogenetic tree. This led to considerable confusion. In 2002, the major research groups came together and formed the Y-Chromosome Consortium (YCC). They published a joint paper that created a single new tree that all agreed to use. Later, a group of citizen scientists with an interest in population genetics and genetic genealogy formed a working group to create an amateur tree aiming at being above all timely. The table below brings together all of these works at the point of the landmark 2002 YCC Tree. This allows a researcher reviewing older published literature to quickly move between nomenclatures.
| YCC 2002/2008 (Shorthand) | (α) | (β) | (γ) | (δ) | (ε) | (ζ) | (η) | YCC 2002 (Longhand) | YCC 2005 (Longhand) | YCC 2008 (Longhand) | YCC 2010r (Longhand) | ISOGG 2006 | ISOGG 2007 | ISOGG 2008 | ISOGG 2009 | ISOGG 2010 | ISOGG 2011 | ISOGG 2012 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| O-M175 | 26 | VII | 1U | 28 | Eu16 | H9 | I | O* | O | O | O | O | O | O | O | O | O | O |
| O-M119 | 26 | VII | 1U | 32 | Eu16 | H9 | H | O1* | O1a | O1a | O1a | O1a | O1a | O1a | O1a | O1a | O1a | O1a |
| O-M101 | 26 | VII | 1U | 32 | Eu16 | H9 | H | O1a | O1a1 | O1a1a | O1a1a | O1a1 | O1a1 | O1a1a | O1a1a | O1a1a | O1a1a | O1a1a |
| O-M50 | 26 | VII | 1U | 32 | Eu16 | H10 | H | O1b | O1a2 | O1a2 | O1a2 | O1a2 | O1a2 | O1a2 | O1a2 | O1a2 | O1a2 | O1a2 |
| O-P31 | 26 | VII | 1U | 33 | Eu16 | H5 | I | O2* | O2 | O2 | O2 | O2 | O2 | O2 | O2 | O2 | O2 | O2 |
| O-M95 | 26 | VII | 1U | 34 | Eu16 | H11 | G | O2a* | O2a | O2a | O2a | O2a | O2a | O2a | O2a | O2a | O2a1 | O2a1 |
| O-M88 | 26 | VII | 1U | 34 | Eu16 | H12 | G | O2a1 | O2a1 | O2a1 | O2a1 | O2a1 | O2a1 | O2a1 | O2a1 | O2a1 | O2a1a | O2a1a |
| O-SRY465 | 20 | VII | 1U | 35 | Eu16 | H5 | I | O2b* | O2b | O2b | O2b | O2b | O2b | O2b | O2b | O2b | O2b | O2b |
| O-47z | 5 | VII | 1U | 26 | Eu16 | H5 | I | O2b1 | O2b1a | O2b1 | O2b1 | O2b1a | O2b1a | O2b1 | O2b1 | O2b1 | O2b1 | O2b1 |
| O-M122 | 26 | VII | 1U | 29 | Eu16 | H6 | L | O3* | O3 | O3 | O3 | O3 | O3 | O3 | O3 | O3 | O3 | O3 |
| O-M121 | 26 | VII | 1U | 29 | Eu16 | H6 | L | O3a | O3a | O3a1 | O3a1 | O3a1 | O3a1 | O3a1 | O3a1 | O3a1 | O3a1a | O3a1a |
| O-M164 | 26 | VII | 1U | 29 | Eu16 | H6 | L | O3b | O3b | O3a2 | O3a2 | O3a2 | O3a2 | O3a2 | O3a2 | O3a2 | O3a1b | O3a1b |
| O-M159 | 13 | VII | 1U | 31 | Eu16 | H6 | L | O3c | O3c | O3a3a | O3a3a | O3a3 | O3a3 | O3a3a | O3a3a | O3a3a | O3a3a | O3a3a |
| O-M7 | 26 | VII | 1U | 29 | Eu16 | H7 | L | O3d* | O3c | O3a3b | O3a3b | O3a4 | O3a4 | O3a3b | O3a3b | O3a3b | O3a2b | O3a2b |
| O-M113 | 26 | VII | 1U | 29 | Eu16 | H7 | L | O3d1 | O3c1 | O3a3b1 | O3a3b1 | - | O3a4a | O3a3b1 | O3a3b1 | O3a3b1 | O3a2b1 | O3a2b1 |
| O-M134 | 26 | VII | 1U | 30 | Eu16 | H8 | L | O3e* | O3d | O3a3c | O3a3c | O3a5 | O3a5 | O3a3c | O3a3c | O3a3c | O3a2c1 | O3a2c1 |
| O-M117 | 26 | VII | 1U | 30 | Eu16 | H8 | L | O3e1* | O3d1 | O3a3c1 | O3a3c1 | O3a5a | O3a5a | O3a3c1 | O3a3c1 | O3a3c1 | O3a2c1a | O3a2c1a |
| O-M162 | 26 | VII | 1U | 30 | Eu16 | H8 | L | O3e1a | O3d1a | O3a3c1a | O3a3c1a | O3a5a1 | O3a5a1 | O3a3c1a | O3a3c1a | O3a3c1a | O3a2c1a1 | O3a2c1a1 |
Original research publications
The following research teams per their publications were represented in the creation of the YCC Tree.
Phylogenetic trees
This phylogenetic tree of haplogroup O subclades is based on the YCC 2008 tree (Karafet 2008) and subsequent published research.
- O1b2 (IMS-JST022454, L272.2, M176/Page63/SRY465, M302, P49, F1942/Page92)
- O1b2a (F1942/Page92)
- O1b2a1 (CTS9259)
- O1b2a1a (F3356)
- O1b2a1a1 (47z, CTS713, CTS11986)
- O1b2a1a2 (F2868, F3110, K4)
- O1b2a1a2a (L682)
- O1b2a1a2b (F940, F1912, F3390)
- O1b2a1a3 (CTS10687, F1800)
- O1b2a1b (CTS562)
- O1b2a1a (F3356)
- O1b2a2 (Page90)
- O1b2a1 (CTS9259)
- O1b2a (F1942/Page92)
Table of frequencies of O-M176
| Population | Frequency | Sample Size | SNPs | Source |
|---|---|---|---|---|
| Korean (Gangweon) | 0.397 | 63 | M176(x47z)=20 47z=5 | Kim 2011 |
| Korean (National Biobank of Korea) | 0.377 | 300 | M176(x47z)=88 47z=25 | Park 2013 |
| South Korea | 0.373 | 75 | M176/P49(x47z)=25 47z=3 | Hammer 2006 |
| Japanese (Shizuoka) | 0.344 | 61 | 47z=13 M176/P49(x47z)=8 | Hammer 2006 |
| Korean (Daejeon) | 0.338 | 133 | M176(x47z)=30 47z=15 | Park 2012 |
| Japanese | 0.335 | 263 | 47z=66 M176/JST022454(x47z)=22 | Nonaka 2007 |
| Korean (Jeju) | 0.322 | 87 | M176(x47z)=20 47z=8 | Kim 2011 |
| Japanese (Kyushu) | 0.321 | 53 | 47z=15 M176/P49(x47z)=2 | Hammer 2006 |
| Korean (Seoul) | 0.316 | 573 | M176(x47z)=125 47z=56 | Park 2012 |
| Korean (Jeolla) | 0.311 | 90 | M176(x47z)=21 47z=7 | Kim 2011 |
| Japanese (Aomori) | 0.308 | 26 | 47z=7 M176/P49(x47z)=1 | Hammer 2006 |
| Korean (Chungcheong) | 0.306 | 72 | M176(x47z)=15 47z=7 | Kim 2011 |
| Japanese (Tokushima) | 0.300 | 70 | 47z=17 M176/P49(x47z)=4 | Hammer 2006 |
| Korean (Gyeongsang) | 0.298 | 84 | M176(x47z)=15 47z=10 | Kim 2011 |
| Japanese | 0.293 | 157 | 47z=38 M176(x47z)=8 | Kim 2011 |
| Korean (Seoul/Gyeonggi) | 0.282 | 110 | M176(x47z)=23 47z=8 | Kim 2011 |
| Korean (PRC) | 0.280 | 25 | M176(x47z)=5 47z=2 | Xue 2006 |
| Korean (Korea) | 0.279 | 43 | M176(x47z)=6 47z=6 | Xue 2006 |
| Japanese | 0.277 | 47 | 47z=11 M176(x47z)=2 | Xue 2006 |
| Manchurians | 0.271 | 48 | 47z=9 M176(x47z)=4 | Jin 2009 |
| Korean (Seoul & Daejeon) | 0.269 | 216 | M176(x47z)=37 47z=21 | Kim 2007 |
| Japanese | 0.262 | 107 | 47z=21 M176(x47z)=7 | Jin 2009 |
| Okinawa | 0.222 | 45 | 47z=5 M176/P49(x47z)=5 | Hammer 2006 |
| Korean | 0.201 | 154 | M176(x47z)=22 47z=9 | Jin 2009 |
| Indonesians | 0.194 | 36 | M176(x47z)=6 47z=1 | Jin 2009 |
| Vietnamese | 0.171 | 41 | M176(x47z)=5 47z=2 | Jin 2009 |
| Indonesia (West) | 0.160 | 25 | M176/P49(x47z)=2 47z=2 | Hammer 2006 |
| Han (Yunnan) | 0.098 | 41 | M176(x47z)=4 | Jin 2009 |
| Vietnamese | 0.083 | 48 | M176(x47z)=2 47z=2 | Kim 2011 |
| Indonesian | 0.081 | 37 | M176(x47z)=3 | Kim 2011 |
| Han (Beijing) | 0.062 | 65 | M176(x47z)=4 | Jin 2009 |
| Micronesia | 0.059 | 17 | M176/P49(x47z)=1 | Hammer 2006 |
| Manchu | 0.057 | 35 | M176(x47z)=2 | Xue 2006 |
| Uriankhai (Mongolia) | 0.050 | 60 | M176=3 | Katoh 2005 |
| Mongol (NE Mongolia) | 0.050 | 20 | M176=1 | DiCristofaro 2013 |
| Hezhe (PRC) | 0.044 | 45 | M176(x47z)=2 | Xue 2006 |
| Vietnam | 0.043 | 70 | 47z=2 M176/P49(x47z)=1 | Hammer 2006 |
| Thai | 0.040 | 50 | 47z=2 | Jin 2009 |
| Manchu | 0.038 | 52 | M176/P49(x47z)=2 | Hammer 2006 |
| Zakhchin (Mongolia) | 0.033 | 60 | M176=2 | Katoh 2005 |
| Manchurian | 0.033 | 30 | M176(x47z)=1 | Kim 2011 |
| Hakka (Taiwan) | 0.029 | 34 | M176=1 | Trejaut 2014 |
| Han (Xi'an) | 0.029 | 34 | M176(x47z)=1 | Kim 2011 |
| Buryat | 0.028 | 36 | M176(x47z)=1 | Kim 2011 |
| Daur | 0.026 | 39 | M176(x47z)=1 | Xue 2006 |
| Thai | 0.025 | 40 | 47z=1 | Kim 2011 |
| Evenk (PRC) | 0.024 | 41 | M176/P49(x47z)=1 | Hammer 2006 |
| Xibe | 0.024 | 41 | M176(x47z)=1 | Xue 2006 |
| Buryat | 0.020 | 50 | M176(x47z)=1 | Jin 2009 |
| Han (Beijing) | 0.020 | 51 | M176(x47z)=1 | Kim 2011 |
| Philippines | 0.014 | 69 | M176(x47z)=1 | Jin 2009 |
| Mongols (Mongolia) | 0.006 | 160 | M176=1 | DiCristofaro 2013 |
| Han (Taiwan) | 0.004 | 258 | M176=1 | Trejaut 2014 |
| Zhuang | 0.000 | 20 | M176/P49=0 | Hammer 2006 |
| Oroqen | 0.000 | 22 | M176/P49=0 | Hammer 2006 |
| Hanoi, Vietnam | 0.000 | 24 | M176=0 | Trejaut 2014 |
| Kalimantan | 0.000 | 25 | M176=0 | Trejaut 2014 |
| Evenk (PRC) | 0.000 | 26 | M176=0 | Xue 2006 |
| Sumatra | 0.000 | 26 | M176=0 | Trejaut 2014 |
| Akha (Thailand) | 0.000 | 27 | M176=0 | Trejaut 2014 |
| Alor | 0.000 | 28 | M176=0 | Karafet 2010 |
| Han (Lanzhou) | 0.000 | 30 | M176=0 | Xue 2006 |
| Even | 0.000 | 31 | M176/P49=0 | Hammer 2006 |
| Oroqen | 0.000 | 31 | M176=0 | Xue 2006 |
| Uyghur (Urumqi) | 0.000 | 31 | M176=0 | Xue 2006 |
| Han (Yili) | 0.000 | 32 | M176=0 | Xue 2006 |
| Malay | 0.000 | 32 | M176/P49=0 | Hammer 2006 |
| Australian aborigines | 0.000 | 33 | M176/P49=0 | Hammer 2006 |
| Qiang | 0.000 | 33 | M176=0 | Xue 2006 |
| Han (Chengdu) | 0.000 | 34 | M176=0 | Xue 2006 |
| Hani (PRC) | 0.000 | 34 | M176=0 | Xue 2006 |
| Li | 0.000 | 34 | M176=0 | Xue 2006 |
| She | 0.000 | 34 | M176=0 | Xue 2006 |
| Buyi | 0.000 | 35 | M176=0 | Xue 2006 |
| Han (Harbin) | 0.000 | 35 | M176=0 | Xue 2006 |
| Han (Meixian) | 0.000 | 35 | M176=0 | Xue 2006 |
| Hui (PRC) | 0.000 | 35 | M176=0 | Xue 2006 |
| Tibetan | 0.000 | 35 | M176=0 | Xue 2006 |
| Yao (Bama) | 0.000 | 35 | M176=0 | Xue 2006 |
| Yao (Liannan) | 0.000 | 35 | M176=0 | Xue 2006 |
| Batak Toba (Sumatra) | 0.000 | 38 | M176=0 | Karafet 2010 |
| Uyghur (Yili) | 0.000 | 39 | M176=0 | Xue 2006 |
| East Indonesia | 0.000 | 40 | M176=0 | Trejaut 2014 |
| Han (Guangdong) | 0.000 | 40 | M176/P49=0 | Hammer 2006 |
| Yi (Butuo, Sichuan) | 0.000 | 43 | M176/P49=0 | Hammer 2006 |
| Northern Han | 0.000 | 44 | M176/P49=0 | Hammer 2006 |
| Khalkh | 0.000 | 45 | M176=0 | Kim 2011 |
| Mongol (Inner Mongolia) | 0.000 | 45 | M176=0 | Xue 2006 |
| Papua New Guinea | 0.000 | 46 | M176/P49=0 | Hammer 2006 |
| Khalkh | 0.000 | 48 | M176=0 | Jin 2009 |
| Philippines | 0.000 | 48 | M176/P49=0 | Hammer 2006 |
| Taiwanese aborigines | 0.000 | 48 | M176/P49=0 | Hammer 2006 |
| Tujia | 0.000 | 49 | M176/P49=0 | Hammer 2006 |
| She | 0.000 | 51 | M176/P49=0 | Hammer 2006 |
| Melanesia | 0.000 | 53 | M176/P49=0 | Hammer 2006 |
| Mandar (Sulawesi) | 0.000 | 54 | M176=0 | Karafet 2010 |
| Han (Fujian) | 0.000 | 55 | M176=0 | Trejaut 2014 |
| Indonesia (East) | 0.000 | 55 | M176/P49=0 | Hammer 2006 |
| Miao | 0.000 | 58 | M176/P49=0 | Hammer 2006 |
| Han (Yunnan) | 0.000 | 60 | M176=0 | Kim 2011 |
| Minnan (Taiwan) | 0.000 | 60 | M176=0 | Trejaut 2014 |
| Nias | 0.000 | 60 | M176=0 | Karafet 2010 |
| Polynesia | 0.000 | 60 | M176/P49=0 | Hammer 2006 |
| Yao | 0.000 | 60 | M176/P49=0 | Hammer 2006 |
| Java | 0.000 | 61 | M176=0 | Karafet 2010 |
| Filipino | 0.000 | 64 | M176=0 | Kim 2011 |
| Mongol (Outer Mongolia) | 0.000 | 65 | M176=0 | Xue 2006 |
| Uyghur | 0.000 | 67 | M176/P49=0 | Hammer 2006 |
| Mentawai | 0.000 | 74 | M176=0 | Karafet 2010 |
| Thailand | 0.000 | 75 | M176=0 | Trejaut 2014 |
| Buryat | 0.000 | 81 | M176/P49=0 | Hammer 2006 |
| Han (Taiwan) | 0.000 | 84 | M176/P49=0 | Hammer 2006 |
| Borneo | 0.000 | 86 | M176=0 | Karafet 2010 |
| Sri Lanka | 0.000 | 91 | M176/P49=0 | Hammer 2006 |
| Lembata | 0.000 | 92 | M176=0 | Karafet 2010 |
| Evenk (Russia) | 0.000 | 95 | M176/P49=0 | Hammer 2006 |
| Altai | 0.000 | 98 | M176/P49=0 | Hammer 2006 |
| Tibet | 0.000 | 105 | M176/P49=0 | Hammer 2006 |
| Java | 0.000 | 141 | M176=0 | Trejaut 2014 |
| Philippines | 0.000 | 146 | M176=0 | Trejaut 2014 |
| Mongolia | 0.000 | 149 | M176/P49=0 | Hammer 2006 |
| Sumba | 0.000 | 350 | M176=0 | Karafet 2010 |
| Taiwan mountain tribes | 0.000 | 355 | M176=0 | Trejaut 2014 |
| Taiwan plains tribes | 0.000 | 370 | M176=0 | Trejaut 2014 |
| Flores | 0.000 | 394 | M176=0 | Karafet 2010 |
| India | 0.000 | 405 | M176/P49=0 | Hammer 2006 |
| Bali | 0.000 | 641 | M176=0 | Karafet 2010 |
See also
Genetics
Y-DNA O subclades
Y-DNA backbone tree
| Phylogenetic tree of human Y-chromosome DNA haplogroups [χ 1][χ 2] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| "Y-chromosomal Adam" | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A00 | A0-T [χ 3] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A0 | A1 [χ 4] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A1a | A1b | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A1b1 | BT | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| B | CT | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DE | CF | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| D | E | C | F | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| F1 | F2 | F3 | GHIJK | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| G | HIJK | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IJK | H | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IJ | K | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| I | J | LT [χ 5] | K2 [χ 6] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| L | T | K2a [χ 7] | K2b [χ 8] | K2c | K2d | K2e [χ 9] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| K-M2313 [χ 10] | K2b1 [χ 11] | P [χ 12] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| NO | S [χ 13] | M [χ 14] | P1 | P2 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| N | O | Q | R | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
References
Footnotes
- ↑ 256/800=32.0% O-M176 in a pool of all Japanese samples of (Xue 2006), (Katoh 2004), (Jin 2009), (Nonaka 2007), (Poznik 2016), and all non-Ainu and non-Okinawan Japanese samples of (Hammer 2006).
- ↑ 202/677=29.8% O-M176 in a pool of all ethnic Korean samples of (Hammer 2006), (Xue 2006), (Katoh 2004), (Kim 2007), and (Park 2013).
- ↑ 30/132=22.7% O-M176 in a pool of all Okinawan data from (Hammer 2006) and (Nonaka 2007)
- ↑ 45/232=19.4% O-M176 in a pool of all Manchu samples of (Karafet 2001), (Jin 2003), (Katoh 2004), and (Xue 2006)
- ↑ 150/628=23.9% O-47z in a pool of all non-Ainu and non-Okinawan Japanese samples of (Jin 2003), (Hammer 2006), (Xue 2006), and (Nonaka 2007)
- ↑ 22/132=16.7% O-47z in a pool of all Okinawan samples of Hammer 2006 and Nonaka 2007
- ↑ 41/519=7.9% O-47z in a pool of all ethnic Korean samples of (Jin 2003), (Hammer 2006), (Xue 2006), and (Kim 2007)
- ↑ 9/135=6.7% O-47z in a pool of all "Manchu" or "Manchurian" samples of (Hammer 2006), (Xue 2006), and (Jin 2009)
- ↑ 41/519=7.9% O-L682 in a pool of all ethnic Korean samples of (Yoo 2014), (Katoh 2004), and (Kim 2011)
- ↑ 22/132=16.7% O-47z in a pool of all Okinawan samples of Hammer 2006 and Nonaka 2007
- ↑ 150/628=23.9% O-47z in a pool of all non-Ainu and non-Okinawan Japanese samples of (Jin 2003), (Hammer 2006), (Xue 2006), and (Nonaka 2007)
- ↑ 9/135=6.7% O-47z in a pool of all "Manchu" or "Manchurian" samples of (Hammer 2006), (Xue 2006), and (Jin 2009)
Works cited
- 1 2 3 4 5 6 YFull Haplogroup YTree v6.01 at 04 January 2018
- 1 2 3 4 5 6 7 8 9 YFull Haplogroup YTree v5.04 at 16 May 2017
- ↑ Kim, Soon-Hee; Kim, Ki-Cheol; Shin, Dong-Jik; et al. "High frequencies of Y-chromosome haplogroup O2b-SRY465 lineages in Korea: a genetic perspective on the peopling of Korea". Investigative Genetics. 2011 (2): 10.
- ↑ Balaresque, Patricia; Poulet, Nicolas; Cussat-Blanc, Sylvain; et al. "Y-chromosome descent clusters and male differential reproductive success: young lineage expansions dominate Asian pastoral nomadic populations". European Journal of Human Genetics. 23: 1413–1422. doi:10.1038/ejhg.2014.285. PMC 4430317. PMID 25585703.
- 1 2 Soon-Hee Kim 2011, High frequencies of Y-chromosome haplogroup O2b-SRY465 lineages in Korea: a genetic perspective on the peopling of Korea
- ↑ Trejaut, Jean A; Poloni, Estella S; Yen, Ju-Chen; et al. (2014). "Taiwan Y-chromosomal DNA variation and its relationship with Island Southeast Asia". BMC Genetics. 15: 77. doi:10.1186/1471-2156-15-77.
- ↑ Huang Dai-Xin, Yang Qing-En, Yin Hui et al., "Genetic Polymorphism of 23 Y Chromosome Biallelic Markers in Wuhan Han Population," HEREDITAS (Beijing) 28 (7) 791~798, 2006
- ↑ Yan, Shi; Chuan-Chao, Wang; Hui, Li; Li, Shi-Lin; Jin, Li; Schurr, Theodore G; Santos, Fabricio R; Quintana-Murci, Lluis; Bertranpetit, Jaume; Comas, David; Tyler-Smith, Chris; Zalloua, Pierre A; Balanovska, Elena; Balanovsky, Oleg; John Mitchell, R; Jin, Li; Soodyall, Himla; Pitchappan, Ramasamy; Cooper, Alan; Matisoo-Smith, Lisa; Royyuru, Ajay K; Platt, Daniel E; Parida, Laxmi; Blue-Smith, Jason; Soria Hernanz, David F; Spencer Wells, R (2011). "An updated tree of Y-chromosome Haplogroup O and revised phylogenetic positions of mutations P164 and PK4". European Journal of Human Genetics. 19 (9): 1013–1015. doi:10.1038/ejhg.2011.64. PMC 3179364. PMID 21505448.
- 1 2 Yali Xue, Tatiana Zerjal, Weidong Bao, Suling Zhu, Qunfang Shu, Jiujin Xu, Ruofu Du, Songbin Fu, Pu Li, Matthew E. Hurles, Huanming Yang, and Chris Tyler-Smith, "Male Demography in East Asia: A North–South Contrast in Human Population Expansion Times." Genetics 172: 2431–2439 (April 2006).
- 1 2 Michael F. Hammer, Tatiana M. Karafet, Hwayong Park, Keiichi Omoto, Shinji Harihara, Mark Stoneking, and Satoshi Horai, "Dual origins of the Japanese: common ground for hunter-gatherer and farmer Y chromosomes." Journal of Human Genetics (2006) 51:47–58. DOI 10.1007/s10038-005-0322-0
- ↑ Wang, C-C; Wang, L-X; Shrestha, R; Zhang, M; Huang, X-Y; et al. (2014). "Genetic Structure of Qiangic Populations Residing in the Western Sichuan Corridor". PLoS ONE. 9 (8): e103772. doi:10.1371/journal.pone.0103772. PMC 4121179. PMID 25090432.
- ↑ Lu Yan (2011), "Genetic Mixture of Populations in Western China." Shanghai: Fudan University, 2011: 1-84. (Doctoral dissertation in Chinese: 陆艳, “中国西部人群的遗传混合”, 上海:复旦大学,2011: 1-84.)
- 1 2 3 So Yeun Kwon, Hwan Young Lee, Eun Young Lee, Woo Ick Yang, and Kyoung-Jin Shin, "Confirmation of Y haplogroup tree topologies with newly suggested Y-SNPs for the C2, O2b and O3a subhaplogroups." Forensic Science International: Genetics 19 (2015) 42–46
- 1 2 G. David Poznik, Yali Xue, Fernando L. Mendez, et al., "Punctuated bursts in human male demography inferred from 1,244 worldwide Y-chromosome sequences." Nature Genetics 2016 June ; 48(6): 593–599. doi:10.1038/ng.3559.
- ↑ Youichi Sato, Toshikatsu Shinka, Ashraf A. Ewis, Aiko Yamauchi, Teruaki Iwamoto, and Yutaka Nakahori, "Overview of genetic variation in the Y chromosome of modern Japanese males." Anthropological Science Vol. 122(3), 131–136, 2014.
- Jin, Han-Jun; Kwak, Kyoung-Don; Hammer, Michael F.; Nakahori, Yutaka; Shinka, Toshikatsu; Lee, Ju-Won; Jin, Feng; Jia, Xuming; et al. (2003). "Y-chromosomal DNA haplogroups and their implications for the dual origins of the Koreans". Human Genetics. 114 (1): 27–35. doi:10.1007/s00439-003-1019-0. PMID 14505036.
- Hammer, Michael F.; Karafet, Tatiana M.; Park, Hwayong; Omoto, Keiichi; Harihara, Shinji; Stoneking, Mark; Horai, Satoshi (2005). "Dual origins of the Japanese: Common ground for hunter-gatherer and farmer Y chromosomes". Journal of Human Genetics. 51 (1): 47–58. doi:10.1007/s10038-005-0322-0. PMID 16328082.
- Jin, Han-Jun; Tyler-Smith, Chris; Kim, Wook (2009). Batzer, Mark A, ed. "The Peopling of Korea Revealed by Analyses of Mitochondrial DNA and Y-Chromosomal Markers". PLoS ONE. 4 (1): e4210. doi:10.1371/journal.pone.0004210. PMC 2615218. PMID 19148289.
- Jin, Han-Jun; Kim, Ki-Cheol; Kim, Wook (2009). "Genetic diversity of two haploid markers in the Udegey population from southeastern Siberia". American Journal of Physical Anthropology. 142: NA. doi:10.1002/ajpa.21232. PMID 19953529.
- Karafet, Tatiana; Xu, Liping; Du, Ruofu; Wang, William; Feng, Shi; Wells, R.S.; Redd, Alan J.; Zegura, Stephen L.; Hammer, Michael F. (2001). "Paternal Population History of East Asia: Sources, Patterns, and Microevolutionary Processes". The American Journal of Human Genetics. 69 (3): 615–28. doi:10.1086/323299. PMC 1235490. PMID 11481588.
- Katoh, Toru; Munkhbat, Batmunkh; Tounai, Kenichi; Mano, Shuhei; Ando, Harue; Oyungerel, Ganjuur; Chae, Gue-Tae; Han, Huun; Jia, Guan-Jun; Tokunaga, Katsushi; Munkhtuvshin, Namid; Tamiya, Gen; Inoko, Hidetoshi (2005). "Genetic features of Mongolian ethnic groups revealed by Y-chromosomal analysis". Gene. 346: 63–70. doi:10.1016/j.gene.2004.10.023. PMID 15716011.
- Kim, Wook; Yoo, Tag-Keun; Kim, Sung-Joo; Shin, Dong-Jik; Tyler-Smith, Chris; Jin, Han-Jun; Kwak, Kyoung-Don; Kim, Eun-Tak; Bae, Yoon-Sun (2007). Blagosklonny, Mikhail, ed. "Lack of Association between Y-Chromosomal Haplogroups and Prostate Cancer in the Korean Population". PLoS ONE. 2 (1): e172. doi:10.1371/journal.pone.0000172. PMC 1766463. PMID 17245448.
- Lu, C.; Zhang, J.; Li, Y.; Xia, Y.; Zhang, F.; Wu, B.; Wu, W.; Ji, G.; et al. (2009). "The b2/b3 subdeletion shows higher risk of spermatogenic failure and higher frequency of complete AZFc deletion than the gr/gr subdeletion in a Chinese population". Human Molecular Genetics. 18 (6): 1122–30. doi:10.1093/hmg/ddn427. PMID 19088127.
- Nonaka, I.; Minaguchi, K.; Takezaki, N. (2007). "Y-chromosomal Binary Haplogroups in the Japanese Population and their Relationship to 16 Y-STR Polymorphisms". Annals of Human Genetics. 71 (4): 480–95. doi:10.1111/j.1469-1809.2006.00343.x. PMID 17274803.
- Tajima, Atsushi; Hayami, Masanori; Tokunaga, Katsushi; Juji, Takeo; Matsuo, Masafumi; Marzuki, Sangkot; Omoto, Keiichi; Horai, Satoshi (2004). "Genetic origins of the Ainu inferred from combined DNA analyses of maternal and paternal lineages". Journal of Human Genetics. 49 (4): 187–93. doi:10.1007/s10038-004-0131-x. PMID 14997363.
- Xue, Y.; Zerjal, T; Bao, W; Zhu, S; Shu, Q; Xu, J; Du, R; Fu, S; et al. (2005). "Male Demography in East Asia: A North-South Contrast in Human Population Expansion Times". Genetics. 172 (4): 2431–9. doi:10.1534/genetics.105.054270. PMC 1456369. PMID 16489223.