RNA-polymerase III
RNA-polymerase III is een RNA-polymerase, die betrokken is bij de productie van onder andere tRNA, siRNA en 5S rRNA en bestaat uit 17 subeenheden met een totale moleculaire massa van ongeveer 0,7 MDa. Het is de grootste RNA-polymerase en komt voor in het cytoplasma van de cel.
Voor het scheiden van de verschillende RNA-polymerasen wordt gebruikgemaakt van DEAE-Sephadex (diethylaminoethylcellulose; een ionenwisselaarhars) bij de ionenuitwisselingschromatografie. Deze techniek scheidt ze van elkaar in de volgorde van de extractie Ι, ΙΙ en ΙΙΙ door de concentratie van ammoniumsulfaat te verhogen. De enzymen werden vernoemd naar de volgorde van extractie RNA-polymerase I, RNA-polymerase II en RNA-polymerase III.[1]
Subeenheden
| Saccharomyces cerevisiae Pol III subeenheden |
Synoniemen | Al of niet in de andere RNA-polymerasen voorkomend |
Menselijk gen | Aantal proteïnogene aminozuren[2] | Menselijke Pol III subeenheden |
|---|---|---|---|---|---|
| RPC1 | RPC155, hRPC155 | III | POLR3A[3] | 1390 | hRPC155, RNA polymerase subunit A |
| RPC2 | FLJ10388, C128 | III | POLR3B[4] | 1133 | RNA polymerase subunit B |
| RPC3 | RPC62 | III | POLR3C[5] | 534 | RNA polymerase subunit C |
| RPC4 | TSBN51 | III | POLR3D[6] | 398 | RNA polymerase subunit D |
| RPC5 | SIN, FLJ10509 | III | POLR3E[7] | 708 | RNA polymerase subunit C5 |
| RPC6 | RPC39 | I, II, III, IV, V | POLR3F[8] | 316 | RNA polymerase F |
| RPC7 | RPC31 | III | POLR3G[9] | 223 | RNA polymerase G |
| RPC8 | KIAA1665 | I, II, III, IV, V | POLR3H[10] | 204 | RNA polymerase H |
| RPC9 | III | POLR3I[11] | 148 | RNA polymerase III subunit RPC9 | |
| RPC10 | III | POLR3J[12] | 108 | RNA polymerases I, II en III subunit RPABC4 | |
| RPC11 | I, II, III, IV, V | POLR3K[13] | 110 | RNA polymerase III subunit K | |
| RPA40 | RPCA1, RPC40, AC40 | I, III | POLR1A[14] | 346 | hRPA40, RNA polymerase I subunit C |
| RPB14 | RPABC1, ABC27 | I, II, III | POLR2E[15] | 210 | hRPB5, RNA polymerase II subunit E |
| RPB6 | RPABC2, ABC23 | I, II, III | POLR2F[16] | 127 | hRPB6, RNA polymerase II subunit F |
| RPB8 | RPABC3, ABC14,5 | I, II, III | POLR2H[17] | 149 | hRPB8, RNA polymerase II subunit H |
| RPB11 | ABC10β, RPABC4 | I, II, III | POLR2K[18] | 70 | hRPB10, RNA polymerase II subunit K |
| RPB12 | ABC10α, RPABC5 | I, II, III | POLR2L[19] | 67 | hRPB12, RNA polymerase II subunit L |
Getranscribeerde RNA's
De typen RNA's, die door RNA-polymerase III getranscribeerd worden, zijn:
- Transfer RNAs[20]
- 5S ribosomaal RNA[20]
- U6 spliceosomaal RNA[20]
- RNase P (Ribonuclease P) en RNase MRP RNA[20]
- 7SL RNA (de RNA-component van het signaalherkenningsdeeltje (SRP))[20]
- Vault RNA's (Vault=vault cytoplasmatisch ribonucleoproteïne)[20]
- Y RNA[20]
- SINE's (SINE = Engels voor kort verspreid kernelement)[20]
- 7SK RNA[20]
- siRNA (small interfering RNA)
- Verscheidene microRNA's[20]
- Verscheidene small nucleolar RNA's (snoRNA)[20]
- Verscheidene genregulerende antisense-RNA's[21]
Bepaling type RNA-polymerase
Alfa-amanitine wordt door zijn werkingsmechanisme ook veelal gebruikt als een stuk gereedschap in wetenschappelijke studies in moleculaire biologie en biologisch onderzoek. Het kan gebruikt worden om te bepalen welke vormen van RNA-polymerase aanwezig zijn. Men test dan de gevoeligheid van de RNA-polymerase in aanwezigheid van alfa-amanitine. RNA-polymerase I, RNA-polymerase IV en RNA-polymerase V zijn ongevoelig, RNA-polymerase II zeer gevoelig en RNA-polymerase III is enigszins gevoelig voor alfa-amanitine.[22]
RNA-polymerase III-transcriptiecyclus
Bij de transcriptie worden drie hoofdstadia onderscheiden:[23]
- Initiatie: de binding van het RNA-polymerasecomplex aan de promotoren van de genen met behulp van transcriptiefactoren.
- Elongatie: de actuele transcriptie van het merendeel van genen in de overeenkomstige RNA-sequentie.
- Terminatie: het stoppen van de RNA-transcriptie en het ontmantelen van het RNA-polymerasecomplex.
RNA-polymerase III werkt samen met de drie transcriptiefactors: TFIIIA, TFIIIB en TFIIIC.
Initiatie
RNA-polymerase III heeft in het algemeen voor de initiatie geen bovenstroomse controle genen nodig, omdat het gebruik maakt van interne controle sequenties in het transcriptiegebied. Soms zijn er wel bovenstroomse controle genen nodig, zoals bij het U6 snRNA-gen dat een bovenstroomse TATA-box heeft, net zoals bij RNA-polymerase II promotoren.
Klasse I
Stadia bij de 5S-rRNA-gen initiatie:
- TFIIIA (Transcriptie Factor voor polymerase III A) bindt aan de intragenetische (liggend in de te transcriberende DNA-sequentie) 5S-rRNA controle sequentie, het C Blok (ook box C genoemd).
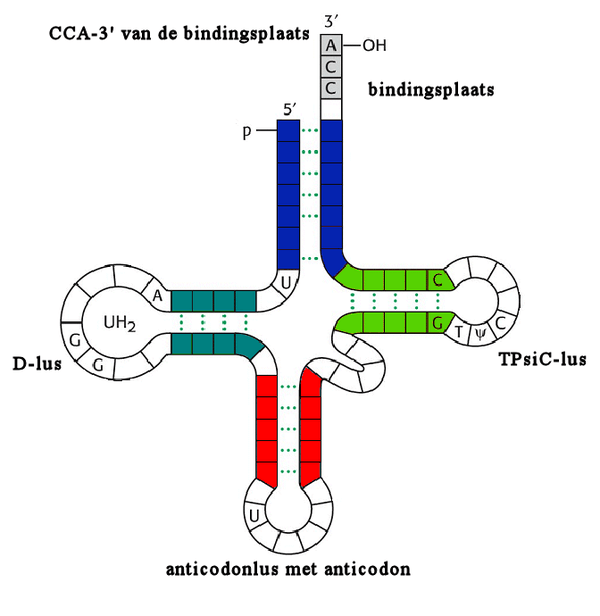
- TFIIIA werkt als een platform dat de A en B blokken vervangt voor de positionering van TFIIIC in een oriëntatie dat rekening houdt met de startplaats van de transcriptie en is vergelijkbaar met het proces bij tRNA genen. De A- en B -blokken vallen samen met de D- en TPsiC-lussen van het tRNA.
- Wanner TFIIIC is gebonden aan het TFIIIA-DNA complex, begint de vorming van TFIIIB, zoals beschreven is bij de tRNA-transcriptie.
Een voorbeeld van een somatisch 5S-rRNA-gen is dat van de klauwkikker (Xenopus laevis) dat drie interne sequenties heeft voor een efficiënte transcriptie: een A blok dat ligt tussen +50 en +64, een B blok tussen +67 en +72 en een C blok tussen +80 en +97.[24]
Klasse II
Stadia bij de tRNA-gen initiatie:
- TFIIIC (Transcriptie Factor voor polymerase III C) bindt aan twee intragenetische controle sequenties, de A- en B-blokken (ook box A en box B genoemd).[25]
- TFIIIC werkt als een vormingsfactor dat het TFIIIB aan het DNA bindt op een plaats ongeveer 26 basenparen bovenstrooms van de startplaats van de transcriptie. TFIIIB (Transcriptioe Factor voor polymerase III B) bestaat uit de drie subeenheden:
- het TBP (TATA Bindings Proteïne),
- het RNA-polymerase II transcriptiefactor TFIIB verwante proteïne, BRF1 (of BRF2 voor transcriptie van de subeenheid van RNA-polymerase III-getranscribeerde genen bij gewervelden) en
- het BDP1.[26]
- TFIIB is de transcriptiefactor dat het RNA-polymerase III vormt op de startplaats van de transcriptie. Wanneer TFIIB gebonden is aan het DNA, is TFIIIC niet langer nodig. TFIIB speelt ook een essentiële rol bij de opening van de promotor.
TFIIB blijft gebonden aan het DNA na de initiatie van de transcriptie door RNA-polymerase III. Hierdoor ontstaat een hoge snelheid van transcriptionele herinitiatie van RNA-polymerase III-getranscribeerde genen.
Klasse III
Stadia bij de U6 snRNA-gen initiatie (alleen onderzocht bij gewervelden):
- SNAPc (SNRNA Activerend Proteïne complex) (ook PBP (CREB-bindingseiwit) en PTF genoemd) bindt aan het PSE (Proximaal Sequentie Element) op een plaats ongeveer 55 basenparen bovenstrooms van de startplaats van de transcriptie. De vorming wordt sterk gestimuleerd door de RNA-polymerase II transcriptiefactoren Oct1 (een Octamer transcriptiefactor bindt aan de "ATTTGCAT"-sequentie} en STAF (Selenocysteïne tRNA-gen Transcriptie Activerings Factor), die binden aan een enhancerachtig DSE (Distaal Sequentie Element) tenminste 200 basenparen bovenstrooms van de startplaats van de transcriptie. Deze factoren en promotor elementen komen gezamenlijk voor tussen de RNA-polymerase II en III transcriptie van snRNA-genen.
- SNAPc zorgt voor de vorming van TFIIB aan de TATA-box op 26 basenparen bovenstrooms van de startplaats van de transcriptie. De aanwezigheid van de TATA-box zorgt voor de transcriptie van het snRNA-gen en niet door RNA-polymerase II.
- De TFIIB voor U6 snRNA transcriptie bevat een kleinere paraloog van BRF1: BRF2.
- TFIIB is de transcriptiefactor voor de vorming van RNA-polymerase III aan de startplaats van de transcriptie.
Elongatie
De RNA-synthese kan worden gestart en de RNA-streng groeit in 5' → 3'-richting. Deze fase wordt elongatie genoemd. Tijdens de elongatie moet telkens het dubbelstrengs-DNA uit elkaar worden gehouden vlak voor de RNA-polymerase. Dit wordt door een transcriptiefactor bewerkstelligd, namelijk TFIIH (transcriptiefactor II met helicase activiteit). Er wordt na de elongatie geen 5'-cap aangebracht, zoals bij de andere polymerasen. Bij U6 snRNA vindt wel monomethylering van een γ-fosfaatrest plaats.
Terminatie
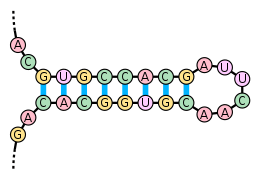
RNA-polymerase III stopt de transcriptie op een klein polyTs (polythymines) gedeelte (5-6). Bij eukaryoten is een haarspeldlus niet nodig, maar wel bij prokaryoten.
Externe link
Bronnen, noten en/of referenties
|