Olenekian
In the geologic timescale, the Olenekian is an age in the Early Triassic epoch; in chronostratigraphy, it is a stage in the Lower Triassic series. It spans the time between 251.2 Ma and 247.2 Ma (million years ago).[2] The Olenekian is sometimes divided into the Smithian and the Spathian subages or substages.[3] The Olenekian follows the Induan and is followed by the Anisian (Middle Triassic).[4]
| System/ Period |
Series/ Epoch |
Stage/ Age |
Age (Ma) | |
|---|---|---|---|---|
| Jurassic | Lower/ Early |
Hettangian | younger | |
| Triassic | Upper/ Late |
Rhaetian | 201.3 | ~208.5 |
| Norian | ~208.5 | ~227 | ||
| Carnian | ~227 | ~237 | ||
| Middle | Ladinian | ~237 | ~242 | |
| Anisian | ~242 | 247.2 | ||
| Lower/ Early |
Olenekian | 247.2 | 251.2 | |
| Induan | 251.2 | 251.902 | ||
| Permian | Lopingian | Changhsingian | older | |
| Subdivision of the Triassic system according to the ICS, as of 2020.[1] | ||||
The Olenekian saw the deposition of a large part of the Buntsandstein in Europe. The Olenekian is roughly coeval with the regional Yongningzhenian stage used in China.

Stratigraphic definitions
The Olenekian stage was introduced into scientific literature by Russian stratigraphers in 1956.[5] The stage is named after Olenëk in Siberia. Before the subdivision in Olenekian and Induan became established, both stages formed the Scythian stage, which has since disappeared from the official timescale.
The base of the Olenekian is at the lowest occurrence of the ammonoids Hedenstroemia or Meekoceras gracilitatis, and of the conodont Neospathodus waageni. It is defined as ending near the lowest occurrences of genera Japonites, Paradanubites, and Paracrochordiceras; and of the conodont Chiosella timorensis. A GSSP (global reference profile for the base) has in 2009 not yet been established.
Olenekian life


Life was still recovering from the severe end-Permian mass extinction. During the Olenekian, the flora changed from lycopod dominated (e.g. Pleuromeia) to gymnosperm and pteridophyte dominated.[6][7] These vegetation changes are due to global changes in temperature and precipitation. Conifers (gymnosperms) were the dominant plants during most of the Mesozoic. Among land vertebrates, the archosaurs - a group of diapsid reptiles encompassing crocodiles, pterosaurs, dinosaurs, and ultimately birds - first evolved from archosauriform ancestors during the Olenekian. This group includes ferocious predators like Erythrosuchus.
In the oceans, Microbial reefs were common during the Early Triassic, possibly due to lack of competition with metazoan reef builders as a result of the extinction.[8] However, transient metazoan reefs reoccurred during the Olenekian wherever permitted by environmental conditions.[9] Ammonoids and conodonts diversified, but both suffered losses during the Smithian-Spathian boundary extinction[10] at the end of the Smithian subage.
Ray-finned fishes largely remained unaffected by the Permian-Triassic extinction event.[11][12] Many genera show a cosmopolitan (worldwide) distribution during the Induan and Olenekian (e.g. Australosomus, Birgeria, Parasemionotidae, Pteronisculus, Ptycholepidae, Saurichthys). This is well exemplified in the Griesbachian (early Induan) aged fish assemblages of the Wordie Creek Formation (East Greenland), the Dienerian (late Induan) aged assemblages of the Sakamena Formation (Madagascar), Candelaria Formation (Nevada, United States), and Mikin Formation (Himachal Pradesh, India), and the Smithian aged assemblages of the Vikinghøgda Formation (Spitsbergen, Norway), Thaynes Formation (western United States), and Helongshan Formation (Anhui, China).
Marine temnospondyl amphibians, such as Aphaneramma or Wantzosaurus show wide geographic ranges during the Induan and Olenekian ages. Their fossils are found in Greenland, Spitsbergen, Pakistan and Madagascar.[13] The first marine reptiles appeared during the Olenekian.[13] Hupehsuchia, Ichthyopterygia and Sauropterygia are among the first marine reptiles to enter the scene (e.g. Cartorhynchus, Chaohusaurus, Utatsusaurus, Hupehsuchus, Grippia, Omphalosaurus, Corosaurus). Sauropterygians and ichthyosaurs ruled the oceans during the Mesozoic Era.
A major extinction event occurred during the Olenekian: the Smithian-Spathian boundary extinction.[14] This event was probably caused by late eruptions of the Siberian Traps and led to extinctions among several groups, especially nektonic and pelagic taxa such as ammonoids and conodonts. One of several exceptionally diverse Early Triassic assemblages, the Paris Biota[15] (from near Paris, southeast Idaho), was deposited in the wake of the Smithian-Spathian boundary extinction. It features at least 7 phyla and 20 distinct metazoan orders, including leptomitid protomonaxonid sponges (previously only known from the Paleozoic), thylacocephalans, crustaceans, nautiloids, ammonoids, coleoids, ophiuroids, crinoids, and vertebrates.[16]

- Gnathorhizids
- Trematosaurids
- Ichthyosauromorphs
- Sauropterygians
- Archosaurs
- Erythrosuchids
- Proterosuchids
- Crurotarsi
Cartilaginous fishes
| Chondrichthyes of the Olenekian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
| Lopingian to Late Cretaceous | Svalbard | A hybodont elasmobranch |
| |
| Carboniferous to Early Triassic | Canada | A chondrichthyan with uncertain affinities, known primarily from their feather-like denticles. | ||
| Lopingian to Late Triassic | United States | A synechodontiform elasmobranch | ||
| Triassic | Svalbard | A hybodont elasmobranch | ||
|
Early Triassic | United States | ||


Ray-finned fishes
| Actinopterygii of the Olenekian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
| Triassic | United States, Svalbard | A non-neopterygian |
| |
| Lopingian to Middle Triassic | Svalbard | A non-neopterygian | ||
| Early Triassic | Greenland, Madagascar | A non-neopterygian | ||
| Early Triassic | Svalbard | A platysiagid | ||
| Early Triassic to Middle Triassic | Svalbard | A non-neopterygian | ||
| Triassic | United States, Svalbard, China | A non-neopterygian | ||

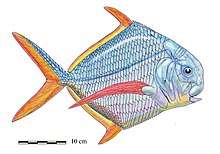

Coelacanths
| Actinistia of the Olenekian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
| Early Triassic | Svalbard | A coelacanth |
| |
| Early Triassic | Svalbard | A coelacanth | ||
| Early Triassic | Canada | A fork-tailed coelacanth | ||
| Early Triassic | Svalbard | A coelacanth | ||
| Early Triassic | Svalbard | A coelacanth | ||
| Early Triassic | Svalbard | A coelacanth | ||

Lungfishes
| Dipnoi of the Olenekian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
| Early Triassic to Eocene | Poland, Russia, Antarctica, Germany | A ceratodontid dipnoan, a close relative of the extant Australian lungfish Neoceratodus. |
| |
| Permian to Early Triassic | Poland, Russia | A gnathorhizid dipnoan | ||
| Early Triassic | Australia | |||
| Early Triassic | Russia | |||


†Temnospondyls
| Temnospondyli of the Olenekian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
| Early Triassic | Svalbard | A trematosaurid amphibian |
| |
| Early Spathian | Poland; Russia | A brachyopid stereospondyl amphibian. | ||
| Early Triassic | Germany | A capitosaurian amphibian | ||
| Early Triassic | Poland; South Africa | A capitosaurian amphibian | ||
| Late Spathian - Early Anisian | Terrigal Formation, Australia | A brachyopid stereospondyl amphibian. | ||
| Germany | A capitosaurian amphibian | |||
| Early Triassic | Germany; Russia | A trematosaurid amphibian | ||


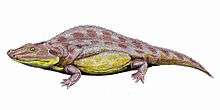

†Chroniosuchians
| Chroniosuchia of the Olenekian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
| Early Triassic | Russia |
| ||
| Lopingian to Early Triassic | Russia, China | |||
|
Early Triassic | Russia | ||
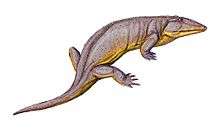

†Procolophonomorphs
| Procolophonomorpha of the Olenekian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
| Early Triassic | Germany, Switzerland | A genus of lizard-shaped parareptile |
| |

Archosauromorphs
| Archosauromorphs of the Olenekian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
| Volgograd Oblast, Russia | The oldest known tanystropheid. |
| ||
| Kolguyev Island, arctic Russia | A basal archosauromorph reptile of controversial classification, various studies have considered it a close relative of Prolacerta, tanystropheids, both, or neither. | |||
| Lipovskaya Formation, Russia | A ctenosauriscid suchian. | |||
| South Africa, China | A proterosuchid that is one of the earliest well-known archosauriforms. It was over 2 meters (6 ft) long and is thought to have behaved like a modern crocodile. Its mouth had two distinct features: the top of its jaw hooked downwards to aid in holding prey, and the upper palate was lined with a row of teeth (a primitive feature lost in later archosaurs). | |||
| Solling Formation, Germany | A ctenosauriscid suchian. Ctenosauriscus is one of the oldest archosaurs known to date. | |||
| Early Triassic | Poland | |||
| 250-237.2 Ma, Olenekian to Ladinian | Omingonde Formation, Namibia South Africa |
The largest erythrosuchid. One of its few distinguishing features other than its size is the smoothness of the margin of the squamosal. In other erythrosuchids, the margin of this bone projects backward from the skull, giving it a hook-like appearance. In Erythrosuchus, the margin is convex and lacks a hook. | ||
| 247.3-245 Ma, Olenekian to Anisian | Russia | A potentially dubious archosauromorph of uncertain affinities. Initially believed to be a proterosuchid archosauriform, a 2016 analysis found that it also shared features with tanystropheids. | ||
| Russia | An erythrosuchid approximately 1.50–2 meters (5–6 ft 8 in) long. | |||
| ~252–247 Ma , Induan to Olenekian | Katberg Formation, East Cape, South Africa | Initially considered to be ancestral to modern lizards, a later study instead found that it shared more similarities with the lineage that would lead to archosaurs. | ||
| 252-250 Ma, Induan to Olenekian | South Africa China |
A proterosuchid that was one of the largest land reptiles during the Early Triassic. It looked somewhat similar to a primitive crocodilian, and shared many of their modern features like long jaws, powerful neck muscles, short legs and a lengthy tail, while possessing several features unique to proterosuchids such as its hook-shaped mouth and long rows of simple cone-shaped teeth. This jaw may have been an adaptation for catching prey, such as Lystrosaurus. | ||
| Russia, | Possibly a rauisuchid. Scythosuchus was between 2 and 3 metres long, and relatively heavily built. | |||
| Induan to Olenekian | Knocklofty Formation, West Hobart, Tasmania | Once believed to be a proterosuchid, this taxon is now believed to have been intermediate between advanced non-archosauriform archosauromorphs such as Prolacerta, and basal archosauriforms such as Proterosuchus. This genus is also notable being one of the most complete Australian Triassic reptiles known. | ||
| Induan to Olenekian | Southern Urals, Russia | Originally classified as a rauisuchid, Tsylmosuchus has more recently been interpreted as an indeterminate archosauriform. Tsylmosuchus occurred throughout the Olenekian age. Some of the strata from which Tsylmosuchus has been found are Induan in age, making it one of the earliest archosaurs. | ||
| latest Olenekian | Yarenskian Gorizont, Komi Republic, Russia | A genus of paracrocodylomorph archosaur, possibly a rauisuchid, and the oldest known archosaur from Russia. | ||
| Heshanggou Formation, China | A ctenosauriscid suchian. Xilousuchus is one of the oldest archosaurs known to date. | |||
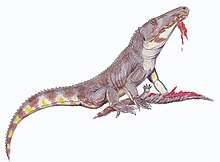
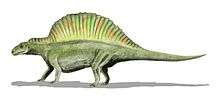
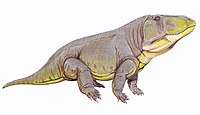
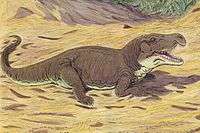


Lepidosauromorphs
| Lepidosauromorpha of the Olenekian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
| Early Triassic | Poland | |||
†Sauropterygia
| Sauropterygians of the Olenekian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
| Late Olenekian | United States, Wyoming | A pistosaur. |
| |
| Olenekian | China | A possible pachypleurosaur. | ||
| Olenekian | China | A possible pachypleurosaur. | ||

†Ichthyosauromorphs
†Hupehsuchians
| †Hupehsuchia of the Olenekian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
| Early Triassic | China | An ichthyosauromorph |
 Unnamed polydactylous hupehsuchian | |
| Early Triassic | China | An ichthyosauromorph | ||
| Early Triassic | China | An ichthyosauromorph | ||
| Early Triassic | China | An ichthyosauromorph | ||
| Early Triassic | China | An ichthyosauromorph | ||

†Ichthyosauriforms
| †Ichthyosauriformes of the Olenekian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
| Early Triassic | China | An early ichthyosauriform |
| |
| Early Triassic | China | An early ichthyosauriform | ||


†Ichthyopterygians
| †Ichthyopterygia of the Olenekian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
| Early Triassic | China |
| ||
| Early Triassic | Canada, Japan, Greenland, China | |||
| Early Triassic to Middle Triassic | Svalbard | An early ichthyosauriform with button-like teeth | ||
| Early Triassic | Svalbard | |||
| Early Triassic | Miyagi Prefecture, Japan; British Columbia, Canada | An early ichthyosauriform | ||




Therapsids
| Therapsids of the Olenekian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
| Kannemeyeria | Burgersdorp Formation, South Africa Ntawere Formation, Zambia Omingonde Formation, Namibia Manda Formation, Tanzania |
A kannemeyeriid that was one of the first representatives of the family, and hence one of the first large herbivores of the Triassic. Although it had a large head, it was lightweight due to the size of the eye sockets and nasal cavity. It also had limb girdles which formed massive plates of bone that helped support its heavily built body. |  | |
| Lystrosaurus | Fremouw Formation, Antarctica | 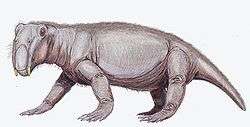 | ||
| Trirachodon | Driekoppen Formation, Free State, South Africa Omingonde Formation, Namibia |
 | ||
References
Notes
- "International Chronostratigraphic Chart" (PDF). International Commission on Stratigraphy. 2020.
- According to Gradstein (2004). Brack et al. (2005) give 251 to 248 Ma
- Tozer E. T. (1965): Lower Triassic stages and ammonoid zones of Arctic Canada: Paper of the Geological Survey of Canada 65:1–14.
- See for a detailed geologic timescale Gradstein et al. (2004)
- Kiparisova & Popov (1956)
- Schneebeli-Hermann, Elke; Kürschner, Wolfram M.; Kerp, Hans; Bomfleur, Benjamin; Hochuli, Peter A.; Bucher, Hugo; Ware, David; Roohi, Ghazala (April 2015). "Vegetation history across the Permian–Triassic boundary in Pakistan (Amb section, Salt Range)". Gondwana Research. 27 (3): 911–924. Bibcode:2015GondR..27..911S. doi:10.1016/j.gr.2013.11.007.
- Goudemand, Nicolas; Romano, Carlo; Leu, Marc; Bucher, Hugo; Trotter, Julie A.; Williams, Ian S. (August 2019). "Dynamic interplay between climate and marine biodiversity upheavals during the early Triassic Smithian -Spathian biotic crisis". Earth-Science Reviews. 195: 169–178. Bibcode:2019ESRv..195..169G. doi:10.1016/j.earscirev.2019.01.013.
- Foster, William J.; Heindel, Katrin; Richoz, Sylvain; Gliwa, Jana; Lehrmann, Daniel J.; Baud, Aymon; Kolar‐Jurkovšek, Tea; Aljinović, Dunja; Jurkovšek, Bogdan; Korn, Dieter; Martindale, Rowan C.; Peckmann, Jörn (20 November 2019). "Suppressed competitive exclusion enabled the proliferation of Permian/Triassic boundary microbialites". The Depositional Record. 6 (1): 62–74. doi:10.1002/dep2.97. PMC 7043383. PMID 32140241.
- Brayard, Arnaud; Vennin, Emmanuelle; Olivier, Nicolas; Bylund, Kevin G.; Jenks, Jim; Stephen, Daniel A.; Bucher, Hugo; Hofmann, Richard; Goudemand, Nicolas; Escarguel, Gilles (18 September 2011). "Transient metazoan reefs in the aftermath of the end-Permian mass extinction". Nature Geoscience. 4 (10): 693–697. Bibcode:2011NatGe...4..693B. doi:10.1038/ngeo1264.
- Galfetti, Thomas; Hochuli, Peter A.; Brayard, Arnaud; Bucher, Hugo; Weissert, Helmut; Vigran, Jorunn Os (2007). "Smithian-Spathian boundary event: Evidence for global climatic change in the wake of the end-Permian biotic crisis". Geology. 35 (4): 291. Bibcode:2007Geo....35..291G. doi:10.1130/G23117A.1.
- Romano, Carlo; Koot, Martha B.; Kogan, Ilja; Brayard, Arnaud; Minikh, Alla V.; Brinkmann, Winand; Bucher, Hugo; Kriwet, Jürgen (February 2016). "Permian-Triassic Osteichthyes (bony fishes): diversity dynamics and body size evolution". Biological Reviews. 91 (1): 106–147. doi:10.1111/brv.12161. PMID 25431138.
- Smithwick, Fiann M.; Stubbs, Thomas L. (2 February 2018). "Phanerozoic survivors: Actinopterygian evolution through the Permo‐Triassic and Triassic‐Jurassic mass extinction events". Evolution. 72 (2): 348–362. doi:10.1111/evo.13421. PMID 29315531.
- Scheyer, Torsten M.; Romano, Carlo; Jenks, Jim; Bucher, Hugo; Farke, Andrew A. (19 March 2014). "Early Triassic Marine Biotic Recovery: The Predators' Perspective". PLOS ONE. 9 (3): e88987. Bibcode:2014PLoSO...988987S. doi:10.1371/journal.pone.0088987. PMC 3960099. PMID 24647136.
- Galfetti, Thomas; Hochuli, Peter A.; Brayard, Arnaud; Bucher, Hugo; Weissert, Helmut; Vigran, Jorunn Os (2007). "Smithian-Spathian boundary event: Evidence for global climatic change in the wake of the end-Permian biotic crisis". Geology. 35 (4): 291. Bibcode:2007Geo....35..291G. doi:10.1130/G23117A.1.
- Brayard, Arnaud; Krumenacker, L. J.; Botting, Joseph P.; Jenks, James F.; Bylund, Kevin G.; Fara, Emmanuel; Vennin, Emmanuelle; Olivier, Nicolas; Goudemand, Nicolas; Saucède, Thomas; Charbonnier, Sylvain; Romano, Carlo; Doguzhaeva, Larisa; Thuy, Ben; Hautmann, Michael; Stephen, Daniel A.; Thomazo, Christophe; Escarguel, Gilles (15 February 2017). "Unexpected Early Triassic marine ecosystem and the rise of the Modern evolutionary fauna". Science Advances. 3 (2): e1602159. Bibcode:2017SciA....3E2159B. doi:10.1126/sciadv.1602159. PMID 28246643.
- Special issue on Paris Biota: https://www.sciencedirect.com/journal/geobios/vol/54
Literature
- Brack, Peter; Rieber, Hans; Nicora, Alda; Mundil, Roland (1 December 2005). "The Global boundary Stratotype Section and Point (GSSP) of the Ladinian Stage (Middle Triassic) at Bagolino (Southern Alps, Northern Italy) and its implications for the Triassic time scale". Episodes. 28 (4): 233–244. doi:10.18814/epiiugs/2005/v28i4/001.
- Gradstein, F.M.; Ogg, J.G. & Smith, A.G.; 2004: A Geologic Time Scale 2004, Cambridge University Press.
- Kiparisova, L.D. & Popov, J.N.; 1956: Расчленение нижнего отдела триасовой системы на ярусы (Subdivision of the lower series of the Triassic System into stages), Doklady Akademii Nauk SSSR 109(4), pp 842–845 (in Russian).
External links
- GeoWhen Database - Olenekian
- Lower Triassic timescale at the website of the subcommission for stratigraphic information of the ICS
- Lower Triassic timescale at the website of Norges Network of offshore records of geology and stratigraphy.