Spinosauridae
| Spinosaurids | |
|---|---|
 | |
| Skeletal reconstruction of Spinosaurus aegyptiacus | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Clade: | Dinosauria |
| Order: | Saurischia |
| Suborder: | Theropoda |
| Superfamily: | †Megalosauroidea |
| Clade: | †Megalosauria |
| Family: | †Spinosauridae Stromer, 1915 |
| Type species | |
| Spinosaurus aegyptiacus Stromer, 1915 | |
| Subgroups | |
| |
| Synonyms | |
Spinosauridae (meaning "spined lizards") is a family of megalosauroidean theropod dinosaurs. The genus Spinosaurus (from which the family, subfamily, and tribe borrow their names) is the largest terrestrial predator known from the fossil record, and likely reached lengths of 15 m (49 ft) or more.[1] Most spinosaurids lived during the Cretaceous Period, and fossils of them have been recovered worldwide, including in Africa, Europe, South America, Asia, and Australia.
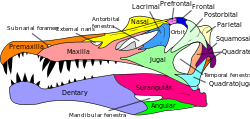
Spinosaurids were large bipedal carnivores with elongated, crocodile-like skulls lined with conical teeth bearing little to no serrations, and small crests on top of their heads. The teeth in the front end of their lower jaws fanned out into a spoon-shaped structure called a rosette, which gave the animal a characteristic look. Their shoulders were robust and prominent, bearing relatively large forelimbs with enlarged claws on the first digit of their hands. Many genera exhibited unusually elongated neural spines, which might have supported sails/humps of skin or fat tissue.
Their diets were composed mostly of aquatic prey, additional fossil evidence indicating they fed opportunistically on other animals like small dinosaurs and pterosaurs. Osteological analyses also suggest a semiaquatic lifestyle for some members of this clade.
Description
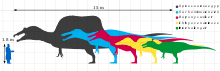
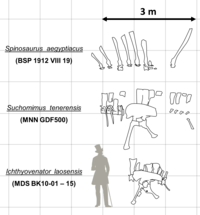
Although reliable size and weight estimates for most taxa are hindered by the lack of good material, all known spinosaurids were large animals.[2] The smallest, Irritator, was between 6 and 8 meters in length and 1 tonne (1.1 short tons) in weight.[3][4] While Ichthyovenator, Baryonyx, and Suchomimus ranged from 7.5 to 11 meters long, and weighing between 1 and 5.2 tonnes (1.2 and 5.7 short tons).[5][6][7] Spinosaurus was the largest, capable of reaching lengths over 15 meters (49 ft)[8] and weighing between 7 and 20.9 tonnes (7.7 and 23.0 short tons),[7][9] making it the largest known terrestrial predator. This consistency in large body size among spinosaurids could have evolved as a byproduct of their preference for semi-aquatic lifestyles, as without the need to compete with other large theropods for food, they would have been enabled to grow to massive lengths.[10]
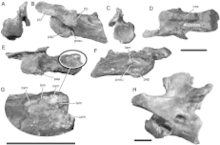
Spinosaurids had relatively large forearms and an enlarged claw on the first digit of the hand.[11] They had hook-shaped coracoids, external nares which are at least behind the teeth of the premaxillae or even further posterior on the skull, a long secondary palate, a terminal rosette of enlarged teeth at the front of the upper and lower jaws, and subconical teeth with either absent or very fine serrations.[11][12] Their skulls often featured low sagittal crests formed from their nasal bones. These crests have been present in Spinosaurus as a ridge-shaped structure, and in Suchomimus and Baryonyx as smaller bumps on top of the skull.[2][13] They have also been seen (to a smaller degree) in Irritator[14] and Cristatusaurus.[15]
Spinosaurus aegyptiacus, the type species for the family and subfamily, is known for the vertebrae with elongated neural spines, some over a meter tall, which have been reconstructed as a sail or hump running down its back.[16] In Ichthyovenator, this sail is a half a meter at its highest and split into two at the sacral vertebrae.[17] Suchomimus also has a low, ridge-like sail over its hips, smaller than that of Spinosaurus.[16] Baryonyx, however, lacks a sail.[18] These structures have had many proposed functions over the years, such as thermoregulation,[19] to aid in swimming,[20] to store energy or insulate the animal, or for display purposes, such as intimidating rivals and predators, or attracting mates.[21][22]
Evolutionary history
Timespan
Spinosaurids are known to exist from as early as the Late Jurassic, through characteristic teeth which were found in Tendaguru, Tanzania, and attributed to Ostafrikasaurus,[23] 15 Million years prior to Siamosaurus. Baryonychines were common, as represented by Baryonyx, which lived during the Barremian of England and Spain. Baryonyx-like teeth are found from the earlier Hauterivian and later Aptian sediments of Spain, as well as the Hauterivian of England, and the Aptian of Niger. The earliest record of spinosaurines is from Africa; they are present in Albian sediments of Tunisia and Algeria, and in Cenomanian sediments of Egypt and Morocco. Spinosaurines are also found in Hauterivian and Aptian-Albian sediments of Thailand, and Southern China. In Africa, baronychines were common in the Aptian, and then replaced by spinosaurines in the Albian and Cenomanian.[24]
Some intermediate specimens extend the known range of spinosaurids past the youngest dates of named taxa. A single baryonychine tooth was found from the mid-Santonian, in the Majiacun Formation of Henan, China.[25] Possible spinosaur remains were also reported from the late Maastrichtian Maevarano Formation.[26]
Localities

Confirmed spinosaurids have been found on every continent except for North America and Antarctica, the first of which was discovered in 1912 at the Bahariya Formation in Egypt and described in 1915 as Spinosaurus aegyptiacus.[22] Africa has shown a great abundance in spinosaurid discoveries,[27] such as in the Kem Kem beds of Morocco, which housed an ecosystem containing many large coexisting predators.[28][29] A fragment of a spinosaurine lower jaw from the Early Cretaceous was also reported from Tunisia, and referred to Spinosaurus.[24]
Spinosaurinae's range has also extended to South America, particularly Brazil, with the discoveries of Irritator, Angaturama, and Oxalaia.[31][32] There was also a fossil tooth in Argentina which has been referred to spinosauridae by Salgado et al.[33] This referral is doubted by Tanaka, who offers Hamadasuchus, a crocodilian, as the most likely animal of origin for these teeth.[34]
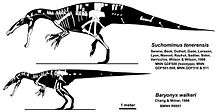
Baryonychines have been found in Africa, with Suchomimus and Cristatusaurus,[24][35][15] as well as in Europe, with Baryonyx and Suchosaurus.[36] Baryonyx-like teeth are also reported from the Ashdown Sands of Sussex, in England, and the Burgos Province, in Spain. A partial skeleton and many fossil teeth indicate spinosaurids were widespread in Asia. As of 2012, three have been named: Ichthyovenator, Siamosaurus and "Sinopliosaurus" fusuiensis.[17][24][25] In 2014, a spinosaurid tooth was discovered in Malaysia; the first dinosaur remains discovered in the country.[37] At la Cantalera-1, a site in the Early Barremanian Blesa Formation in Treul, Spain, two types of spinosaurid teeth were found, and they were assigned, tentatively, as indeterminate spinosaurine and baryonychine taxa.[38]
An intermediate spinosaurid was discovered in the Early Cretaceous Eumeralla Formation, Australia.[39] It is known from a single 4 cm long partial cervical vertebra, designated NMV P221081. It is missing most of the neural arch. The specimen is from a juvenile estimated to be about 2 to 3 meters long (6-9 ft). Out of all spinosaurs it most closely resembles Baryonyx.[40]
Classification

The family Spinosauridae was named by Ernst Stromer in 1915 to include the single genus Spinosaurus. The clade was expanded as more close relatives of Spinosaurus were uncovered. The first cladistic definition of Spinosauridae was provided by Paul Sereno in 1998 (as "All spinosauroids closer to Spinosaurus than to Torvosaurus").[41]
Taxonomy
Traditionally, Spinosauridae is divided into two subfamilies: Spinosaurinae, which contains the genera Icthyovenator, Irritator, Oxalaia, Sigilmassasaurus and Spinosaurus, is marked by unserrated, straight teeth, and external nares which are further back on the skull than in Baryonychinae.[41][42] And Baryonychinae, which contains the genera Baryonyx and Suchomimus,[24] is marked by serrated, slightly curved teeth, smaller size, and more teeth in the lower jaw behind the terminal rosette than in spinosaurines.[41][42] Others, such as Siamosaurus, may belong to either Baryonychinae or Spinosaurinae, but are too incompletely known to be assigned with confidence.[24] Siamosaurus was classified as a spinosaurine in 2018, but the results are provisional and not entirely conclusive.[43]
Phylogeny
The subfamily Spinosaurinae was named by Sereno in 1998, and defined by Holtz et al. (2004) as all taxa closer to Spinosaurus aegyptiacus than to Baryonyx walkeri. And the subfamily Baryonychinae was named by Charig & Milner in 1986. They erected both the subfamily and the family Baryonychinae for the newly discovered Baryonyx, before it was referred to the Spinosauridae. Their subfamily was defined by Holtz et al. in 2004, as the complementary clade of all taxa closer to Baryonyx walkeri than to Spinosaurus aegyptiacus. Examinations by Marcos Sales and Cesar Schultz et al. (2017) indicate that the South American spinosaurids Angaturama, Irritator, and Oxalaia were intermediate between Baronychinae and Spinosaurinae based on their craniodental features and cladistic analysis. This indicates that Baryonychinae may in fact be non-monophyletic. Their cladogram can be seen below.[44]
| Spinosauridae |
| ||||||||||||||||||||||||
The next cladogram displays an analysis of Tetanurae simplified to show only Spinosauridae from Allain et al. (2012):[45]
| Spinosauridae |
| ||||||||||||||||||||||||
The 2018 phylogenetic analysis by Arden et al., which included many unnamed taxa, resolved Baryonychinae as monophyletic, and also coined the new term Spinosaurini for the clade of Sigilmassasaurus and Spinosaurus.[43]
| Spinosauridae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||
Paleobiology
Diet and feeding

Spinosaurid teeth resemble those of crocodiles, which are used for piercing and holding prey. Therefore, teeth with small or no serrations, such as in spinosaurids, were not good for cutting or ripping into flesh but instead helped to ensure a strong grip on a struggling prey animal.[46] Spinosaur jaws were likened by Vullo et al. to those of the pike conger eel, in what they hypothesized was convergent evolution for aquatic feeding.[47] Both kinds of animals have some teeth in the end of the upper and lower jaws that are larger than the others and an area of the upper jaw with smaller teeth, creating a gap into which the enlarged teeth of the lower jaw fit, with the full structure called a terminal rosette.[47]
Spinosaurids have in the past often been considered mainly fish-eaters (piscivores), based on comparisons of their jaws with those of modern crocodilians.[42] Rayfield and colleagues, in 2007, conducted biomechanical studies on the skull of the European spinosaurid Baryonyx, which has a long, laterally compressed skull, comparing it to gharial (long, narrow, tubular) and alligator (flat and wide) skulls.[48] They found that the structure of baryonychine jaws converged on that of gharials, in that the two taxa showed similar response patterns to stress from simulated feeding loads, and did so with and without the presence of a (simulated) secondary palate. The gharial, exemplar of a long, narrow, and tubular snout, is a fish specialist. However, this snout anatomy doesn’t preclude other options for the spinosaurids. While the gharial is the most extreme example and a fish specialist, and Australian freshwater crocodiles (Crocodylus johnstoni), which have similarly shaped skulls to gharials, also specialize more on fish than sympatric, broad snouted crocodiles. And are opportunistic feeders which eat all manner of small aquatic prey, including insects and crustaceans.[48] Thus, their aptly shaped snouts correlate with fish-eating, this is consistent with hypotheses of this diet for spinosaurids, in particular baryonychines, but it does not indicate that they were solely piscivorous.

A further study by Cuff and Rayfield (2013) on the skulls of Spinosaurus and Baryonyx did not recover similarities in the skulls of Baryonyx and the gharial that the previous study did. Baryonyx had, in models where the size difference of the skulls was corrected for, greater resistance to torsion and dorsoventral bending than both Spinosaurus and the gharial, while both spinosaurids were inferior to the gharial, alligator, and slender-snouted crocodile in resisting torsion and medio-lateral bending.[49] When the results from the modeling were not scaled according to size, then both spinosaurids performed better than all the crocodilians in resistance to bending and torsion, due to their larger size.[49] Thus, Cuff and Rayfield suggest that the skulls are not efficiently built to deal well with relatively large, struggling prey, but that the spinosaurids may overcome prey simply by their size advantage, and not skull build[49] Sues and colleagues studied the construction of the spinosaurid skull, and concluded that their mode of feeding was to use extremely quick, powerful strikes to seize small prey items using their jaws, whilst employing the powerful neck muscles in rapid up-and-down motion. Due to the narrow snout, vigorous side-to-side motion of the skull during prey capture is unlikely.[46] Based the size and positions of their nostrils, Sales & Schultz (2017) suggested that Spinosaurus possessed a greater reliance on its sense of smell and had a more piscivorous lifestyle than Irritator and baryonychines.[44]
Ontogeny
Juvenile spinosaurid fossils are exceedingly rare and almost unknown. However, an ungual phalanx measuring 21 mm (0.83 in) belonging to a very young Spinosaurus indicates that Spinosaurus and probably by extent other spinosaurids, may have developed their semiaquatic adaptations at birth or while at a very young age and maintained these adaptations throughout their lives. The specimen, found in 1999 and described by Simone Maganuco and Cristiano Dal Sasso et al. in 2018, is believed to have come from a very small juvenile measuring 1.78 m (5.8 ft), making said specimen the smallest known example of a spinosaurid currently described. [50][51]
Paleoecology

Habitat preference
A 2010 publication by Romain Amiot and colleagues found that oxygen isotope ratios of spinosaurid bones indicates semiaquatic lifestyles. Isotope ratios from teeth from Baryonyx, Irritator, Siamosaurus, and Spinosaurus were compared with isotopic compositions from contemporaneous theropods, turtles, and crocodilians. The study found that, among theropods, spinosaurid isotope ratios were closer to those of turtles and crocodilians. Siamosaurus specimens tended to have the largest difference from the ratios of other theropods, and Spinosaurus tended to have the least difference. The authors concluded that spinosaurids, like modern crocodilians and hippopotamuses, spent much of their daily lives in water. The authors also suggested that semiaquatic habits and piscivory in spinosaurids can explain how spinosaurids coexisted with other large theropods: by feeding on different prey items and living in different habitats, the different types of theropods would have been out of direct competition.[29]
In 2018, an analysis was conducted on the partial tibia of an indeterminate spinosaurine from the early Albian, the bone was from a sub-adult between 7–13 m (22–42 ft) in length still growing moderately fast before its death. This specimen (LPP-PV-0042) was found in the Araripe Basin of Brazil and taken to the University of San Carlos for a CT Scan, where it revealed osteosclerosis (high bone density).[52] This condition had previously only been observed in Spinosaurus, as a possible way of controlling its buoyancy.[53] The presence of this condition on the leg fragment showed that semi-aquatic adaptations in spinosaurids were already present at least 10 million years before Spinosaurus aegyptiacus appeared. According to the Phylogenetic bracketing method, this high bone density might have been present in all spinosaurines.[52]
Feeding

Direct fossil evidence shows that spinosaurids fed on fish as well as a variety of other small to medium-sized animals, including small dinosaurs. Baryonyx was found with scales of the prehistoric fish, Scheenstia, in its body cavity, and these were abraded, hypothetically by gastric juices. Bones of a young Iguanodon, also abraded, were found alongside this specimen. If these represent Baryonyx’s meal, Baryonyx was, whether in this case a hunter, or a scavenger, an eater of more diverse fare than fish.[54][55][56] Moreover, there is a documented example of a spinosaurid having eaten a pterosaur, as spinosaurid teeth were found embedded within the fossil vertebrae of one found in the Santana Formation of Brazil. This may represent a predation event, but Buffetaut et al. consider it more likely that the spinosaurid scavenged the pterosaur carcass after its death.[57] In the Sao Khua Formation of Thailand, isolated tooth crowns from Siamosaurus have been found in association with sauropod remains, indicating possible predation or scavenging.[58]
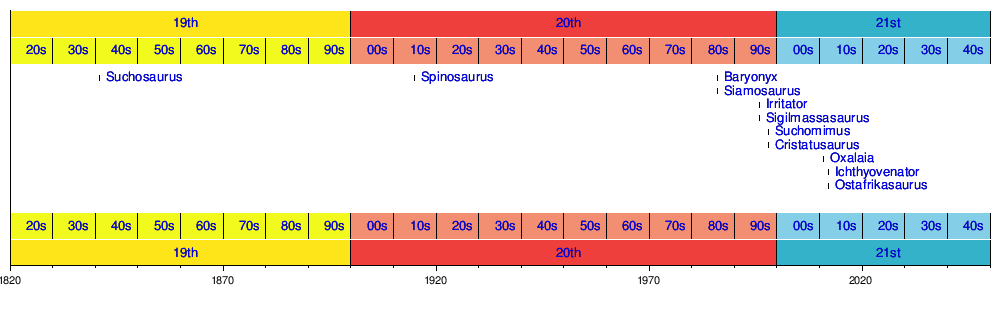
Timeline of genera

Timeline of genera descriptions
References
- ↑ Ibrahim, Nizar; Sereno, Paul C.; Dal Sasso, Cristiano; Maganuco, Simone; Fabri, Matteo; Martill, David M.; Zouhri, Samir; Myhrvold, Nathan; Lurino, Dawid A. (2014). "Semiaquatic adaptations in a giant predatory dinosaur". Science. 345 (6204): 1613–6. Bibcode:2014Sci...345.1613I. doi:10.1126/science.1258750. PMID 25213375. Supplementary Information
- 1 2 HONE, David William Elliott; HOLTZ, Thomas Richard (2017). "A Century of Spinosaurs - A Review and Revision of the Spinosauridae with Comments on Their Ecology". Acta Geologica Sinica - English Edition. 91 (3): 1120–1132. doi:10.1111/1755-6724.13328. ISSN 1000-9515.
- ↑ Dixon, Dougal (2009). The Ultimate Guide to Dinosaurs. Ticktock Books. ISBN 9781846969881.
- ↑ Holtz, Thomas R. Jr. (2011) Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages, Winter 2010 Appendix.
- ↑ S.,, Paul, Gregory. The Princeton field guide to dinosaurs (2nd ed.). Princeton, N.J. ISBN 9781400883141. OCLC 954055249.
- ↑ Holtz, Thomas R. Jr. (2011) Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages, Winter 2010 Appendix.
- 1 2 Therrien, François; Henderson, Donald M. (2007-03-12). "My theropod is bigger than yours … or not: estimating body size from skull length in theropods". Journal of Vertebrate Paleontology. 27 (1): 108–115. doi:10.1671/0272-4634(2007)27[108:mtibty]2.0.co;2. ISSN 0272-4634.
- ↑ Ibrahim, Nizar; Sereno, Paul C.; Dal Sasso, Cristiano; Maganuco, Simone; Fabri, Matteo; Martill, David M.; Zouhri, Samir; Myhrvold, Nathan; Lurino, Dawid A. (2014). "Semiaquatic adaptations in a giant predatory dinosaur". Science. 345 (6204): 1613–6. Bibcode:2014Sci...345.1613I. doi:10.1126/science.1258750. PMID 25213375. Supplementary Information
- ↑ dal Sasso, C.; Maganuco, S.; Buffetaut, E.; Mendez, M.A. (2005). "New information on the skull of the enigmatic theropod Spinosaurus, with remarks on its sizes and affinities" (Submitted manuscript). Journal of Vertebrate Paleontology. 25 (4): 888–896. doi:10.1671/0272-4634(2005)025[0888:NIOTSO]2.0.CO;2. ISSN 0272-4634.
- ↑ Aureliano, Tito; Ghilardi, Aline M.; Buck, Pedro V.; Fabbri, Matteo; Samathi, Adun; Delcourt, Rafael; Fernandes, Marcelo A.; Sander, Martin (3 May 2018). "Semi-aquatic adaptations in a spinosaur from the Lower Cretaceous of Brazil". Cretaceous Research. doi:10.1016/j.cretres.2018.04.024. ISSN 0195-6671.
- 1 2 Sereno, Paul C., Allison L. Beck, Didier B. Dutheil, Boubacar Gado, Hans C. E. Larsson, Gabrielle H. Lyon, Jonathan D. Marcot, et al. 1998. “A Long-Snouted Predatory Dinosaur from Africa and the Evolution of Spinosaurids.” Science 282 (5392): 1298–1302. doi:10.1126/science.282.5392.1298.
- ↑ Rayfield, Emily J. 2011. “Structural Performance of Tetanuran Theropod Skulls, with Emphasis on the Megalosauridae, Spinosauridae and Carcharodontosauridae.” Special Papers in Palaeontology 86 (November). https://www.researchgate.net/publication/250916680_Structural_performance_of_tetanuran_theropod_skulls_with_emphasis_on_the_Megalosauridae_Spinosauridae_and_Carcharodontosauridae.
- ↑ Charig, A. J.; Milner, A. C. (1997). "Baryonyx walkeri, a fish-eating dinosaur from the Wealden of Surrey". Bulletin of the Natural History Museum of London. 53: 11–70.
- ↑ Sues, H. D.; Frey, E.; Martill, D. M.; Scott, D. M. (2002). "Irritator challengeri, a spinosaurid (Dinosauria: Theropoda) from the Lower Cretaceous of Brazil". Journal of Vertebrate Paleontology. 22 (3): 535–547. doi:10.1671/0272-4634(2002)022[0535:ICASDT]2.0.CO;2.
- 1 2 Taquet, P. and Russell, D.A. (1998). "New data on spinosaurid dinosaurs from the Early Cretaceous of the Sahara". Comptes Rendus de l'Académie des Sciences à Paris, Sciences de la Terre et des Planètes 327: 347-353
- 1 2 Hecht, Jeff. 1998. “Fish Swam in Fear.” New Scientist. November 21. https://www.newscientist.com/article/mg16021610-300-fish-swam-in-fear/.
- 1 2 Allain, R.; Xaisanavong, T.; Richir, P.; Khentavong, B. (2012). "The first definitive Asian spinosaurid (Dinosauria: Theropoda) from the early cretaceous of Laos". Naturwissenschaften. 99 (5): 369–377. Bibcode:2012NW.....99..369A. doi:10.1007/s00114-012-0911-7. PMID 22528021.
- ↑ Charig, A. J.; Milner, A. C. (1997). "Baryonyx walkeri, a fish-eating dinosaur from the Wealden of Surrey". Bulletin of the Natural History Museum of London. 53: 11–70.
- ↑ Halstead, L.B. (1975). The Evolution and Ecology of the Dinosaurs. London: Eurobook Limited. pp. 1–116. ISBN 0-85654-018-8.
- ↑ "Gimsa, J., Sleigh, R., Gimsa, U., (2015) : "The riddle of Spinosaurus aegyptiacus ' dorsal sail". University of Rostock, Chair for Biophysics, Gertrudenstr. 11A, 18057 Rostock, Germany".
- ↑ Bailey, J.B. (1997). "Neural spine elongation in dinosaurs: sailbacks or buffalo-backs?". Journal of Paleontology. 71 (6): 1124–1146. JSTOR 1306608.
- 1 2 Stromer, E. (1915). "Ergebnisse der Forschungsreisen Prof. E. Stromers in den Wüsten Ägyptens. II. Wirbeltier-Reste der Baharije-Stufe (unterstes Cenoman). 3. Das Original des Theropoden Spinosaurus aegyptiacus nov. gen., nov. spec". Abhandlungen der Königlich Bayerischen Akademie der Wissenschaften, Mathematisch-physikalische Klasse (in German). 28 (3): 1–32.
- ↑ Buffetaut, Eric. 2008. “Spinosaurid Teeth from the Late Jurassic of Tendaguru, Tanzania, with Remarks on the Evolutionary and Biogeographical History of the Spinosauridae.” https://www.academia.edu/3101178/Spinosaurid_teeth_from_the_Late_Jurassic_of_Tendaguru_Tanzania_with_remarks_on_the_evolutionary_and_biogeographical_history_of_the_Spinosauridae
- 1 2 3 4 5 6 Buffetaut, Eric, and Mohamed Ouaja. 2002. “A New Specimen of Spinosaurus (Dinosauria, Theropoda) from the Lower Cretaceous of Tunisia, with Remarks on the Evolutionary History of the Spinosauridae.” Bulletin de La Société Géologique de France 173 (5): 415–21. doi:10.2113/173.5.415.
- 1 2 Hone, Dave, Xing Xu, Deyou Wang, and Vertebrata PalAsiatica. 2010. “A Probable Baryonychine (Theropoda: Spinosauridae) Tooth from the Upper Cretaceous of Henan Province, China (PDF Download Available).” ResearchGate. January. https://www.researchgate.net/publication/271967379_A_probable_Baryonychine_Theropoda_Spinosauridae_tooth_from_the_Upper_Cretaceous_of_Henan_Province_China.
- ↑ Weishampel, David B.; Barrett, Paul M.; Coria, Rodolfo A.; Le Loueff, Jean; Xu Xing; Zhao Xijin; Sahni, Ashok; Gomani, Elizabeth M. P.; Noto, Christopher N. (2004). "Dinosaur distribution". In Weishampel, David B.; Dodson, Peter; Osmólska, Halszka. The Dinosauria (2nd ed.). Berkeley: University of California Press. p. 624. ISBN 0-520-24209-2.
- ↑ Benyoucef, Madani, Emilie Läng, Lionel Cavin, Kaddour Mebarki, Mohammed Adaci, and Mustapha Bensalah. 2015. “Overabundance of Piscivorous Dinosaurs (Theropoda: Spinosauridae) in the Mid-Cretaceous of North Africa: The Algerian Dilemma.” Cretaceous Research 55 (July): 44–55. doi:10.1016/j.cretres.2015.02.002.
- ↑ Hendrickx, Christophe, Octávio Mateus, and Eric Buffetaut. 2016. “Morphofunctional Analysis of the Quadrate of Spinosauridae (Dinosauria: Theropoda) and the Presence of Spinosaurus and a Second Spinosaurine Taxon in the Cenomanian of North Africa.” PLOS ONE 11 (1): e0144695. doi:10.1371/journal.pone.0144695.
- 1 2 Amiot, R.; Buffetaut, E.; Lécuyer, C.; Wang, X.; Boudad, L.; Ding, Z.; Fourel, F.; Hutt, S.; Martineau, F.; Medeiros, A.; Mo, J.; Simon, L.; Suteethorn, V.; Sweetman, S.; Tong, H.; Zhang, F.; Zhou, Z. (2010). "Oxygen isotope evidence for semi-aquatic habits among spinosaurid theropods". Geology. 38 (2): 139–142. Bibcode:2010Geo....38..139A. doi:10.1130/G30402.1.
- ↑ Farke, Andrew A.; Benson, Roger B. J.; Rich, Thomas H.; Vickers-Rich, Patricia; Hall, Mike (2012). "Theropod Fauna from Southern Australia Indicates High Polar Diversity and Climate-Driven Dinosaur Provinciality". PLoS ONE. 7 (5): e37122. Bibcode:2012PLoSO...737122B. doi:10.1371/journal.pone.0037122. ISSN 1932-6203.
- ↑ Buffetaut, Eric, David Martill, and François Escuillié. 2004. “Pterosaurs as Part of a Spinosaur Diet.” Nature 430. doi:10.1038/430033a.
- ↑ Kellner, Alexander W.A.; Sergio A.K. Azevedeo; Elaine B. Machado; Luciana B. Carvalho; Deise D.R. Henriques (2011). "A new dinosaur (Theropoda, Spinosauridae) from the Cretaceous (Cenomanian) Alcântara Formation, Cajual Island, Brazil" (PDF). Anais da Academia Brasileira de Ciências. 83 (1): 99–108. doi:10.1590/S0001-37652011000100006. ISSN 0001-3765.
- ↑ Salgado, Leonardo, José I. Canudo, Alberto C. Garrido, José I. Ruiz-Omeñaca, Rodolfo A. García, Marcelo S. de la Fuente, José L. Barco, and Raúl Bollati. 2009. “Upper Cretaceous Vertebrates from El Anfiteatro Area, Río Negro, Patagonia, Argentina.” Cretaceous Research 30 (3): 767–84. doi:10.1016/j.cretres.2009.01.001.
- ↑ Tanaka, Gengo. 2017. “Fine Sculptures on a Tooth of Spinosaurus (Dinosauria, Theropoda) from Morocco.” Bulletin of Gunma …. Accessed May 30. https://www.academia.edu/1300482/Fine_sculptures_on_a_tooth_of_Spinosaurus_Dinosauria_Theropoda_from_Morocco.
- ↑ Sereno, P.C.; Beck, A.L.; Dutheil, D.B.; Gado, B.; Larsson, H.C.E.; Lyon, G.H.; Marcot, J.D.; Rauhut, O.W.M.; Sadleir, R.W.; Sidor, C.A.; Varricchio, D.D.; Wilson, G.P; Wilson, J.A. (1998). "A long-snouted predatory dinosaur from Africa and the evolution of spinosaurids". Science. 282 (5392): 1298–1302. Bibcode:1998Sci...282.1298S. doi:10.1126/science.282.5392.1298. PMID 9812890. Retrieved 2013-03-19.
- ↑ Mateus, O.; Araújo, R.; Natário, C.; Castanhinha, R. (2011). "A new specimen of the theropod dinosaur Baryonyx from the early Cretaceous of Portugal and taxonomic validity of Suchosaurus" (PDF). Zootaxa. 2827: 54–68.
- ↑ ResearchSEA (2014). "First discovery of dinosaur fossils in Malaysia". ScienceDaily.
- ↑ Alonso, Antonio, and José Ignacio Canudo. 2016. “On the Spinosaurid Theropod Teeth from the Early Barremian (Early Cretaceous) Blesa Formation (Spain).” Historical Biology 28 (6): 823–34. doi:10.1080/08912963.2015.1036751.
- ↑ "Australian 'Spinosaur' unearthed". Australian Geographic. Retrieved 2018-04-15.
- ↑ Barrett, P. M.; Benson, R. B. J.; Rich, T. H.; Vickers-Rich, P. (2011). "First spinosaurid dinosaur from Australia and the cosmopolitanism of Cretaceous dinosaur faunas". Biology Letters. 7 (6): 933–936. doi:10.1098/rsbl.2011.0466. ISSN 1744-9561.
- 1 2 3 Sereno, Paul C., Allison L. Beck, Didier B. Dutheil, Boubacar Gado, Hans C. E. Larsson, Gabrielle H. Lyon, Jonathan D. Marcot, et al. 1998. “A Long-Snouted Predatory Dinosaur from Africa and the Evolution of Spinosaurids.” Science 282 (5392): 1298–1302. doi:10.1126/science.282.5392.1298.
- 1 2 3 Rayfield, Emily J. 2011. “Structural Performance of Tetanuran Theropod Skulls, with Emphasis on the Megalosauridae, Spinosauridae and Carcharodontosauridae.” Special Papers in Palaeontology 86 (November). https://www.researchgate.net/publication/250916680_Structural_performance_of_tetanuran_theropod_skulls_with_emphasis_on_the_Megalosauridae_Spinosauridae_and_Carcharodontosauridae.
- 1 2 Arden, T.M.S.; Klein, C.G.; Zouhri, S.; Longrich, N.R. (2018). "Aquatic adaptation in the skull of carnivorous dinosaurs (Theropoda: Spinosauridae) and the evolution of aquatic habits in Spinosaurus". Cretaceous Research. In Press. doi:10.1016/j.cretres.2018.06.013.
- 1 2 Sales, M.A.F.; Schultz, C.L. (2017). "Spinosaur taxonomy and evolution of craniodental features: Evidence from Brazil". PLoS ONE. 12 (11): e0187070. Bibcode:2017PLoSO..1287070S. doi:10.1371/journal.pone.0187070.
- ↑ Allain, R.; Xaisanavong, T.; Richir, P.; Khentavong, B. (2012). "The first definitive Asian spinosaurid (Dinosauria: Theropoda) from the early cretaceous of Laos". Naturwissenschaften. 99 (5): 369–377. Bibcode:2012NW.....99..369A. doi:10.1007/s00114-012-0911-7. PMID 22528021.
- 1 2 Sues, Hans-Dieter, Eberhard Frey, David M. Martill, and Diane M. Scott. 2002. “Irritator Challengeri, a Spinosaurid (Dinosauria: Theropoda) from the Lower Cretaceous of Brazil.” Journal of Vertebrate Paleontology 22 (3): 535–47. doi:10.1671/0272-4634(2002)022[0535:ICASDT]2.0.CO;2.
- 1 2 Vullo, R.; Allain, R.; Cavin, L. (2016). "Convergent evolution of jaws between spinosaurid dinosaurs and pike conger eels". Acta Palaeontologica Polonica. 61. doi:10.4202/app.00284.2016.
- 1 2 Rayfield, Emily J., Angela C. Milner, Viet Bui Xuan, and Philippe G. Young. 2007. “Functional Morphology of Spinosaur ‘crocodile-Mimic’ Dinosaurs.” Journal of Vertebrate Paleontology 27 (4): 892–901. doi:10.1671/0272-4634(2007)27[892:FMOSCD]2.0.CO;2.
- 1 2 3 Cuff, Andrew R., and Emily J. Rayfield. 2013. “Feeding Mechanics in Spinosaurid Theropods and Extant Crocodilians.” PLOS ONE 8 (5): e65295. doi:10.1371/journal.pone.0065295.
- ↑ https://www.sciencedaily.com/releases/2018/05/180530132947.htm
- ↑ http://www.sci-news.com/paleontology/smallest-spinosaurus-06052.html
- 1 2 Aureliano, Tito; Ghilardi, Aline M.; Buck, Pedro V.; Fabbri, Matteo; Samathi, Adun; Delcourt, Rafael; Fernandes, Marcelo A.; Sander, Martin (3 May 2018). "Semi-aquatic adaptations in a spinosaur from the Lower Cretaceous of Brazil". Cretaceous Research. doi:10.1016/j.cretres.2018.04.024. ISSN 0195-6671.
- ↑ Ibrahim, N.; Sereno, P. C.; Dal Sasso, C.; Maganuco, S.; Fabbri, M.; Martill, D. M.; Zouhri, S.; Myhrvold, N.; Iurino, D. A. (2014). "Semiaquatic adaptations in a giant predatory dinosaur". Science. 345 (6204): 1613–1616. doi:10.1126/science.1258750. PMID 25213375.
- ↑ Rayfield, Emily J. 2011. “Structural Performance of Tetanuran Theropod Skulls, with Emphasis on the Megalosauridae, Spinosauridae and Carcharodontosauridae.” Special Papers in Palaeontology 86 (November). https://www.researchgate.net/publication/250916680_Structural_performance_of_tetanuran_theropod_skulls_with_emphasis_on_the_Megalosauridae_Spinosauridae_and_Carcharodontosauridae.
- ↑ Sues, Hans-Dieter, Eberhard Frey, David M. Martill, and Diane M. Scott. 2002. “Irritator Challengeri, a Spinosaurid (Dinosauria: Theropoda) from the Lower Cretaceous of Brazil.” Journal of Vertebrate Paleontology 22 (3): 535–47. doi:10.1671/0272-4634(2002)022[0535:ICASDT]2.0.CO;2.
- ↑ Charig, A. J.; Milner, A. C. (1997). "Baryonyx walkeri, a fish-eating dinosaur from the Wealden of Surrey". Bulletin of the Natural History Museum of London. 53: 11–70.
- ↑ Buffetaut, Eric, David Martill, and François Escuillié. 2004. “Pterosaurs as Part of a Spinosaur Diet.” Nature 430. doi:10.1038/430033a.
- ↑ Sales, Marcos A. F.; Lacerda, Marcel B.; Horn, Bruno L. D.; Oliveira, Isabel A. P. de; Schultz, Cesar L. (2016-02-01). "The "χ" of the Matter: Testing the Relationship between Paleoenvironments and Three Theropod Clades". PLOS ONE. 11 (2): e0147031. Bibcode:2016PLoSO..1147031S. doi:10.1371/journal.pone.0147031. ISSN 1932-6203. PMC 4734717. PMID 26829315.
External links
- Spinosauridae on the Theropod Database
| |||||||||||||||||||||
| Piatnitzkysauridae | 
.png) | ||||||||||||||||||||
| Megalosauria |
| ||||||||||||||||||||
.jpg)