Irritator
| Irritator | |
|---|---|
 | |
| Reconstructed skeleton at the National Museum of Nature and Science, Tokyo | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Clade: | Dinosauria |
| Order: | Saurischia |
| Suborder: | Theropoda |
| Family: | †Spinosauridae |
| Subfamily: | †Spinosaurinae |
| Genus: | †Irritator Martill et al. 1996 |
| Species | |
| |
| Synonyms | |
| |
Irritator is a genus of theropod dinosaur that lived in what is now Brazil during the Early Cretaceous Period (Albian), about 110 million years ago. It was first described in 1996 from a nearly complete skull found in the Romualdo Formation, originally identified as a member of the Santana Formation, and later elevated to a separate formation; belonging to the Santana Group in the Araripe Basin. Fossil dealers acquired the skull and illegaly sold it, but not before artificially lengthening it; this required many hours of restoration by paleontologists, who chose the genus name Irritator to express their frustration. The epithet of the type species Irritator challengeri honors Professor Challenger, a character in Arthur Conan Doyle's novels.
Irritator's holotype specimen remains the most complete preserved skull discovered for a spinosaurid. It had a long, shallow snout and slender jaws, which were lined with straight and unserrated conical teeth. A sagittal crest ran lengthwise atop the head, serving as an attachment site for powerful neck muscles. Irritator's nostrils were positioned far back on the head, and a rigid secondary palate on the roof of its mouth would have strengthened the jaw when feeding. A generalist diet, like that of today's crocodilians, has been suggested for Irritator; in which it would have mainly nourished itself on fish and any other small prey animals that it could catch. Fossil evidence is known of an individual that ate a pterosaur, possibly from hunting or scavenging it. Irritator was between 6 and 8 metres (20 and 26 ft) in length, and weighed around 1 tonne (1.1 short tons), making it one of the smallest spinosaurs known.
The first dinosaur described from the Araripe Basin, Irritator's unprepared holotype skull was initially mistaken for that of an early pterosaur, and later a maniraptoran dinosaur closely related to the feathered ornithomimids and troodontids. Later research identified the animal as belonging to the Spinosauridae, a family of large and likely semiaquatic predators. The genus is regarded today as potentially synonymous with Angaturama limai, known from a snout tip that was described from the same stratigraphic units of the Araripe Basin where Irritator was found. Spinosaurid skeletal material is known from the Romualdo Formation, but to what genus it belongs to is uncertain. The formation seldom yields dinosaur fossils; most of the unit's known vertebrate fauna consists largely of pterosaurs and fish, many of which are known from a variety of well-preserved fossils.
History of discovery

Near the town of Santana do Cariri in eastern Brazil, commercial fossil-poachers excavated a chalk concretion containing the rear of a large skull with lower jaws, which were flattened somewhat sideways. This fossil was acquired by dealers who illegally sold it[1] — the fossil trade is since 1942 prohibited by law in Brazil[2] — to Rupert Wild of the State Museum of Natural History Stuttgart.[1] At the time it was assumed to be the skull of a giant basal pterosaur, since the Chapada do Araripe region is famous for its copious amount of pterosaur finds, and the German museum often bought such pieces. As it promised to be a unique discovery of singular importance, German and British pterosaur experts were contacted to study the exemplar. A paper describing it as a pterosaur had already been submitted for publication, when the authors were quickly disabused of the notion it had been a flying reptile by the peer review, in which it was pointed out that the fossil belonged to a theropod dinosaur.[3]
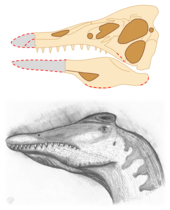
The skull (designated SMNS 58022) became the holotype specimen of the new genus Irritator in 1996, when it was first scientifically described by by paleontologists David M. Martill, Arthur R. I. Cruickshank, Eberhard Frey, Philip G. Small and Malcolm Clarke.[1] The right side was well preserved, while the left sustained extensive damage during collection. Some of the skull's hindmost upper surface had eroded away, and the lower jaw lacked its front end; owing to breakage during fossilization. The front of the rostrum (snout) was also missing, and since there were no signs of erosion, it most probably broke off during or after collection of the fossil. Evident corrosion on certain bones indicates acid preparation had been attempted, and a vertical fracture was present across the middle of the skull, which had apparently been sealed with car body filler. Parts of the specimen were already cracked due to being part of a septarian concretion, but the poor preparation by the collectors further extended the damage.[8] The skull was severely obscured beneath plaster added by the fossil traders in hopes of making it look more complete and valuable. The buyers were unaware of the modifications to the illegally collected specimen until it was sent to universities in the UK for CT scan imaging, which revealed that the collectors had tried to reconstruct the skull by grafting parts of the maxilla (main upper jaw bone) onto the front of the rostrum.[1] Martill and his team, in 1996, wrote that the generic name Irritator came "from irritation, the feeling the authors felt (understated here) when discovering that the snout had been artificially elongated."[1] The type species, Irritator challengeri, honored Professor Challenger; a character in Arthur Conan Doyle's The Lost World.[1][9]
When Martill and colleagues first described Irritator, the holotype was still extensively encased in calcareous rock matrix. Diane Scott from the University of Toronto assumed the task of fully cleaning out the skull, allowing for a detailed redescription in 2002. Written by Hans-Dieter Sues, Frey, Martill, and Scott, this inspection of the now fully prepared specimen negated many of Martill and colleagues' original observations, which were based on misinterpretations of the damaged skull. For example: the estimated length of the complete skull was 24 cm (9.4 in) shorter than previously proposed, various new skull bones were identified, and what was originally thought to be a prominent head crest proved to be an unattached, indeterminate bone fragment. As in the previous study, Sues and colleagues regarded the African genus Spinosaurus as the most similar taxon to Irritator, on account of them sharing many dental features; including a mostly straight conical crown, a thin enamel layer, well-defined edges with no serrations, and lengthwise fluting. These similarities were enough for the authors to suggest a possible junior synonymy of Irritator to Spinosaurus, noting however, that more overlapping skull material was needed for further diagnosis.[8] Later research maintained separation of the two taxa.[10][11][12]
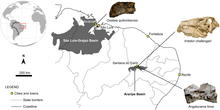
Irritator is the first dinosaur described from the Araripe Basin and although its exact point of origin is unknown, the holotype skull and its accompanying matrix have been assigned to a rock of the then defined Romualdo Member,[1] which is composed almost entirely of limestone.[13] This classification was confirmed by microfossils of the ostracod Pattersoncypris, and fish scales from the ichthyodectid Cladocyclus; both found in the Romualdo Member. Questioning of local fossil dealers resulted in the identification of the discovery site near the village of Buxexé close to Santana Do Cariri at the flank of the Chapada do Araripe, at a height of approximately 650 metres (2,130 ft). Since rock from the Romualdo Member is indeed exposed there, and the holotype has the same color and texture as those rocks, this discovery site is regarded as very probable for the fossil.[8] In 2011, another Brazilian spinosaur, Oxalaia quilombensis, was named and described from the more northwestern Alcântara Formation. This species, known only from an isolated snout tip and upper jaw fragment, lived during the Cenomanian; around six to nine million years after Irritator.[14]
Besides skull elements and isolated teeth, the Romualdo Formation has also yielded postcranial remains from spinosaurids,[15] many of which are hitherto undescribed. Only spinosaurines have been recovered in the Araripe, with no baryonychines reported as of yet.[16] In 2004, parts of a spinal column were unearthed at the formation. Brazilian paleontologists Jonathas Bittencourt and Alexander Kellner assigned these, due to their structure, to the Spinosauridae. It is uncertain whether this specimen, cataloged under the number MN 4743-V, can be referred to Irritator or Angaturama. Given that the last two are based on skull material, direct comparisons cannot be made.[17] In 2007, Elaine Machado and Kellner tentatively assigned a rib fragment (MN 7021-V) to the Spinosauridae.[16] The most complete spinosaur (and theropod) skeleton retrieved from the Romualdo is MN 4819-V, which comprises a largely intact pelvis, a dorsal (back) and caudal (tail) vertebra, five sacral (hip) vertebrae, elements of the right hand, and a partial right tibia and femur. The specimen was referred to the Spinosauridae due to its tall sacral neural spines and the enlarged condition of the recurved manual ungual (claw). This specimen was first mentioned by Kellner in a 2001 abstract, but was not described until 2010 in an as-of-yet unpublished master's thesis by Machado.[15][16][17] In 2018, Tito Aureliano and his team described LPP-PV-0042, part of a left tibia from a particularly large individual.[16] Like many fossils from the Araripe Basin, most of the spinosaurid material from the Romualdo Formation was collected for use in the illegal fossil trade. As such, most of the specimens lack precise geological data on the circumstances of their discovery.[18][16]
Synonymy with Angaturama
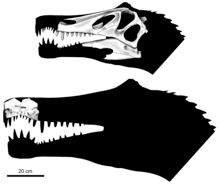
Angaturama limai, from the same time and place as Irritator challengeri, was described by Kellner and Diogenes Campos in February 1996 on the basis of an isolated premaxilla from the Romualdo Formation. The generic name is in reference to Angaturama, a protective spirit in the aboriginal Tupi Indian culture of Brazil, and the specific name alludes to paleontologist Murilo R. de Lima, who informed Kellner of the specimen in 1991. The remains of Angaturama were diagnosed by very strong lateral compression of the snout, and a thin sagittal crest (complete shape unknown) on top of the premaxilla.[19] Subsequent research led to discovery of the various postcranial spinosaur specimens from the Romualdo Formation, which were used as the basis for the creation of a replica skeleton, later mounted for exhibit at the Federal University-owned Museu Nacional do Rio de Janeiro (National Museum of Brazil).[20] The skeleton depicted the animal carrying an anhanguerid pterosaur in its jaws. It was the centerpiece of the "Dinossauros no Sertão" (Dinosaurs of Sertão) exhibit, which opened to the public in March 2009, becoming the first large carnivorous Brazilian dinosaur to be put on display.[21][22] Some of the original postcranial elements (including the fossil pelvis and sacral vertebra) were presented alongside the mount, which was portrayed in a saltwater lagoon environment.[21][23]
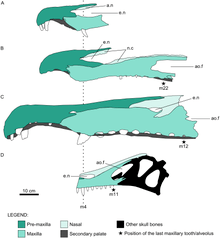
In 1997, British paleontologists Alan J. Charig and Angela C. Milner considered it likely that Angaturama was a junior synonym of Irritator, since both genera had retracted nostrils, long jaws, and characteristic spinosaurid dentition.[24] Paul Sereno and colleagues in 1998 agreed with this possibility, additionally noting that the holotype of Angaturama seems to complete that of Irritator (meaning that they could belong to the same specimen).[25] This conclusion was supported by various authors, including Éric Buffetaut and Mohamed Ouaja in 2002,[12] and Cristiano Dal Sasso and colleagues in 2005.[26] Sues and colleagues followed suit in their redescription of Irritator, stating that both holotypes are equally as narrow, and share transversely round teeth with defined yet unserrated edges. Angaturama's premaxillary saggital crest may also correspond with that of Irritator's nasal bones.[8] Some objection, however, has been raised to these assertions of synonymy. Kellner and Campos in 2000 and Machado and Kellner in 2005 both expressed the opinion that the fossils come from two different genera, and that the holotype of Angaturama was clearly more laterally flattened than Irritator.[3][27] A review of both fossils by Marcos Sales and Cesar Schultz in 2017 noted that the specimens also differ in other aspects of their preservation: the Irritator specimen is brighter in color, and is affected by a vertical crack while the Angaturama specimen bears many cavities; and the damage to the teeth of Irritator's holotype is much less severe. They also identified a possible point of overlap, the third left maxillary tooth, and observed that the skull of Angaturama may have been larger than that of Irritator, based on the proportions of the closely related genus Baryonyx. Nevertheless, although the two specimens evidently do not belong to the same individual, Sales and Schultz noted that synonymy at the genus level would need to be verified by more extensively overlapping remains. If Angaturama and Irritator are actually regarded as a member of the same genus, Irritator challengeri would be the valid scientific name under rules of priority.[15]
Description
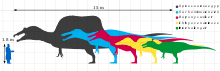
Even by maximal size estimates, Irritator was smaller than other known spinosaurids. In 2010, Gregory S. Paul calculated its length at 7.5 meters (25 ft) and weight at 1 tonne (1.1 short tons).[28] Thomas R. Holtz Jr. in 2011 published a higher estimate of 8 meters (26 ft), with a weight between 0.9 to 3.6 tonnes (0.99 to 3.97 short tons).[29][30] Earlier estimates in 2009 by Dougal Dixon were lower at 6 meters (20 ft) long and 2 metres (6.6 ft) high.[31] In 2018, the reconstructions from the study by Sales and Schultz were scaled by Aureliano and colleagues, providing a length of 6.5 meters (21 ft) for Irritator's holotype (specimen SMNS 58022), and 8.3 meters (27 ft) for Angaturama's holotype (USP GP/2T).[16] Aureliano and colleagues also estimated a total body length of roughly 10 meters (33 ft) for the spinosaurine tibia fragment from the Romualdo (LPP-PV-0042). Bone histology was used to determine that this individual was a subadult undergoing fairly rapid growth before time of death; thus, the fully mature animal might have been even larger.[16] The partial spinosaurine skeleton MN 4819-V represented a moderately-sized specimen, estimated by Machado in 2010 at 5 to 6 meters (16 to 20 ft) in length.[32] Many elements from this specimen were incorporated into the skeletal mount in the National Museum of Brazil, which had a length of 6 meters (20 ft) and a height of 2 metres (6.6 ft).[20][21]
Anatomy of the holotype
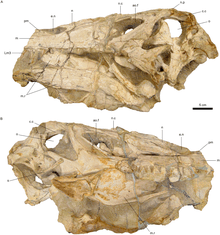
The holotype skull, although heavily damaged in certain places, is largely complete; missing only the tip of the snout (including part of the maxilla and most of the premaxilla) and the front of the mandible. The preserved skull is 16.5 cm (6.5 in) tall and 10 cm (3.9 in) wide, its full length has been estimated at roughly 60 cm (24 in); based on comparisons with Baryonyx. Irritator's skull was long, narrow, and somewhat triangular in cross section. The braincase was inclined backwards, and deeper than it was long. Extending from it was an elongate and narrow snout, with both sides relatively flat and slightly angled towards the skull midline.[8] The nares (nostril openings) were oval and, as in all spinosaurids, positioned higher and further back on the skull than in typical theropods. Irritator's nares were smaller than in Suchomimus and Baryonyx, but larger than those of Spinosaurus.[8][10] The opening behind the orbit (eye socket), the lateral temporal fenestra, was very large; while the antorbital fenestra, located in front of the eyes, was long and ellipse-shaped. The orbit itself was deep and wider at the top (where the eyeball was placed) than the bottom. The lacrimal bone separated the orbit from the antorbital fenestra, forming the upper and lower rear margins of the latter with two processes that enclosed a 40 degree angle; similar to Baryonyx, in which it enclosed 35 degrees. The prefrontal bones were large and sturdy, while the thinner frontal bones, situated behind them, were smooth and concave on top; both of these bones formed the upper rim of the orbit. Irritator's holotype is unique in that it is one of the few non-avian dinosaur specimens found with a preserved columella (equivalent to the stapes in mammals).[8]

A thin sagittal crest, constructed from the elongate nasal bones, ran up the skull midline before stopping just above the eye in a slightly flattened bulge.[8] Although the complete shape and height of this structure is unknown in Irritator, these head crests were commonplace in spinosaurids, having possibly served a display function when alive.[33] The preserved part of Irritator's crest lacks the vertical ridges seen in that of Spinosaurus.[12] Like other members of its family, Irritator possessed a long and bony structure on the roof of its mouth called a secondary palate; a feature observed in extant crocodilians, but absent in most theropod dinosaurs.[8][34] The back of the lower jaw was deep, its rear upper surface consisting mainly of the large surangular bone, which articulated with the shallower angular bone below it. The mandibular fenestra, a sideways-facing opening in the lower jaw, was oval and comparatively large. The dentary (tooth-bearing bone of the mandible) is unknown in Irritator, save for a possible remnant at the front of the surangular. Only the rear ends of the paired premaxillae (frontmost snout bone) remain intact, forming the front upper and lower borders of the nostril openings. As in other spinosaurids, the maxillary bones formed a long and low branch that extended under the nostrils, which separated the premaxillae from the nasals. Some of the holotype's skull bones had not yet fully co-ossified (fused), indicating that the specimen belonged to a subadult individual.[8]

Irritator had straight or only faintly recurved conical teeth, bearing sharply defined but unserrated edges. Lengthwise flutes (ridges) were present on its tooth crowns, a common dental trait among spinosaurids.[8][10] Both sides of Irritator's teeth were fluted, as in Spinosaurus; while Baryonyx exhibited them only on the lingual (inward facing) side of its teeth. Irritator's teeth were circular in cross section, as opposed to the laterally flattened condition of most theropod teeth. The enamel (first layer) was thin, with a finely wrinkled texture also observed in Baryonyx. The left and right maxillae of Irritator preserve nine teeth, although the former's tooth crowns are more intact, and there are traces of a tenth tooth in the rock matrix. The teeth were deeply inserted, and widely spaced towards the front of the maxilla. The first and second preserved maxillary teeth were the largest, at 32 mm (1.3 in) and 40 mm (1.6 in) in crown length respectively.[8] The seven remaining teeth became progressively smaller towards the rear, one of the last ones measuring 6 mm (0.24 in) in estimated crown length. CT scans performed on the specimen revealed replacement teeth on either side of the upper jaw, whose roots ran deep into the maxillae and converged close to the midline, nearly reaching the top of the skull.[8][1] Based on comparisons with relatives, the maxillae were probably lined with a total of 11 teeth each, similar to the number of 12 teeth in MSNM V4047, an upper jaw referred to Spinosaurus.[10] At the time of this Irritator specimen's death, the hindmost tooth of the left maxilla was still being replaced, and only the tip of it was visible.[8]
Anatomy of the "Angaturama" holotype

The holotype of Angaturama consists only of the front part of the upper jaw, comprising the tip of the premaxillae and the frontmost end of the maxillae; kept today under the number USP GP/2T-5 in the University of São Paulo. The specimen measures 19.2 cm (7.6 in) in height and 11 cm (4.3 in) in length, with the width of the palatal region being 4 to 5 mm (0.16 to 0.20 in). The suture between the maxilla and premaxilla was jagged at its front and straightened out towards the rear. The lip region was curved at the sixth premaxillary tooth, and the front of the snout was expanded, forming the spoon-shaped terminal rosette characteristic of spinosaurids. This concave underside of the premaxillae would have complemented a convex and also enlarged tip of the mandible.[19] The premaxillae connected with each other on the bottom to form Angaturama's secondary palate, which was also partially contributed to by two processes extending from the maxillae.[10] The snout was strongly compressed laterally, and the premaxillae gently tapered towards the top to form a tall sagittal crest 1 to 2 mm (0.039 to 0.079 in) in thickness. This crest was larger, and extended further forwards on the snout than in other known spinosaurids. The frontmost upper border of the premaxilla had a small bulge that overhung the base of the crest. This bulge was apparently damaged on its upper surface, indicating that the top of the crest may have extended even farther over and forwards from that point. The front of Angaturama's snout hence had a vertically straight or concave margin, atypical from the more smoothly-sloping snouts of other spinosaurs.[10][19]
In the premaxilla, a broken-off tooth with partial tooth crown was recovered. The strongly extended and straight teeth with unserrated conical crowns, which measured 6 to 40 mm (0.24 to 1.57 in) in length, were singly embedded. This indicates continuous tooth replacement where new teeth were pushed up between the old ones. Judging by the tooth sockets, altogether the premaxilla had seven teeth; the third tooth being the largest. The frontmost three teeth of the maxilla were also preserved. The premaxillary teeth increased in size from the first to third, shrink from the third to the sixth, and enlarge again from the sixth premaxillary to tenth maxillary positions. A 16 cm (6.3 in) diastema (gap in tooth row) was present between the last premaxillary and first maxillary tooth.[19]
Classification
Martill and his team originally believed Irritator to be an unusual new genus of maniraptoran dinosaur in the clade Bullatosauria (now Maniraptoriformes[35]), as a close relative to the feathered ornithomimosaurs and troodontids. Given that its long snout, dental morphology, and assumed fin-shaped crest were features unknown in "other" maniraptorans, the researchers erected the new family Irritatoridae within the clade, based on these traits.[1] They recognized Irritator's affinities with Spinosaurus, in that they both had similarly shaped and unserrated teeth; noting, however, that the latter's holotype mandible would not conform with their reconstruction of Irritator's upper jaw, and that other non-avian dinosaurs like Compsognathus and Ornitholestes also bore no serrations on some or all of their teeth.[1] In 1996, Kellner found no evidence supporting Martill's referral of Irritator to the maniraptora, and instead resolved the genus as a spinosaurid; a conclusion agreed upon by later studies.[8] Irritator was then assigned to the family Baryonychidae along with Angaturama, Baryonyx, Suchomimus, and Spinosaurus by Oliver Rauhut in 2003.[36] Thomas Holtz and colleagues in 2004 considered the Baryonychidae synonymous with Spinosauridae, and placed these genera within that family.[37] Most later revisions have upheld these classifications.[33][11]
In 1998, Paul Sereno defined two subfamilies within the Spinosauridae on the basis of craniodental characteristics. They were Spinosaurinae, in which he placed Spinosaurus and Irritator; and Baryonychinae, to which he assigned Baryonyx, Suchomimus, and Cristatusaurus. Irritator was separated from baryonychines by its unserrated, straighter, and more widely spaced teeth.[38] In 2005, a reconstruction of Irritator's skull by Dal Sasso and colleagues assumed the external nares as being located above the middle of the maxillary tooth row; more posteriorly than in baryonychines, but less so than in Spinosaurus.[12] However, Sales and Schultz's later identification of the third left maxillary tooth's position, placed Irritator's external nares in a comparable position to those of Baryonyx and Suchomimus. The more forward nostril placement was typically considered characteristic of baryonychines. Nevertheless, Irritator also bore unserrated teeth, a trait associated with spinosaurines. Sales and Schultz noted that the South American spinosaurids Irritator, Angaturama, and Oxalaia might represent intermediate forms between Baryonychinae and Spinosaurinae, and that with further research and evidence, the former could eventually be rendered a paraphyletic group.[15] Irritator is further distinguished from Baryonyx, Suchomimus, and Cristatusaurus by having almost half as many teeth in the maxilla. It is also considered distinct from Spinosaurus due to its comparatively larger and more forwardly positioned nostril openings, which unlike in Spinosaurus, are also formed by the premaxilla. The narrow saggital crest, which ends in a knob-like process above the frontals, is another autapomorphy (distinguishing feature) of Irritator from other spinosaurs.[15]
Although the snout of Angaturama is generally narrower than in baryonychines, part of the extreme sideways flattening of the holotype may be due to taphonomy (changes during decay and fossilization); given that the specimen appears partly crushed and broken, with some of the preserved teeth having been sectioned off along their length. Therefore, Angaturama's only valid autapomorphy is its sagittal crest; which extends further forwards on the rostrum and is more exaggerated than in other known spinosaurid skulls.[15][39]
The following cladogram shows a 2009 analysis of the Megalosauroidea.[40]
| Megalosauroidea |
| ||||||||||||||||||||||||
The next cladogram follows the results of the cladistic study by Sales and Schultz in 2017.[15]
| Spinosauridae |
| ||||||||||||||||||||||||
Paleobiology

In 1996, Martill and colleagues theorized that Irritator, with its elongated snout and unserrated conical teeth, likely had a piscivorous (fish-eating) diet, like the pterosaurs found in large numbers at the Romualdo Formation.[1] Although much of the holotype's morphology turned out to be greatly different, later studies supported these observations.[15][33] Spinosaurids had very narrow jaws with relatively homogeneous pointed teeth.[33] This arrangement is found particularly in crocodiles such as the false gharial.[34] The long conical teeth, which do not possess serrated edges, were suitable for the grabbing and holding of prey. They differed from teeth of other theropods, which seemed geared towards tearing or cutting off seized body parts.[33]

The nostrils of Irritator were shifted far to the rear of the skull; this, along with the secondary palate, which separated the animal's nasal passages from the inside of its mouth, made respiration possible even if the majority of the jaw was underwater or held prey. In particular, the sagittal crest of Irritator is an indication for pronounced neck musculature, which would have been necessary in order to pull the jaw closed quickly against water resistance and withdraw the head fast.[8] In 2015, Serjoscha W. Evers and colleagues found evidence for similar adaptations in the African spinosaur Sigilmassasaurus. The neck vertebra of this genus have a heavily furrowed undersurface, which is consistent with the attachment of powerful neck muscles for use in fishing or rapidly snatching small prey; a trait also observed in extant crocodilians and birds.[41] In 2002, Sues and colleagues pointed out, however, that there would be no reason to assume that the Spinosauridae specialized completely in fishing. They stress rather that this head morphology indicates a generalistic feeding, particularly on small prey animals. In fact, portions of a young Iguanodon, a terrestrial herbivore, were found inside the fossil skeleton of one Baryonyx.[8] Darren Naish and colleagues in 2004 supported the theory that Irritator hunted both aquatic and terrestrial animals as a generalist within the coastal area and in addition probably searched for carrion.[42] A tooth belonging to Irritator was discovered still inserted into the fossil neck vertebral column of an ornithocheirid pterosaur, likely with a wingspan of 3.3 m (11 ft). This indicates that Irritator ate pterosaurs as well. Although it is not known if it actively hunted these animals, or simply scavenged the remains.[43][44]
Paleoecology
Irritator and Angaturama are known from the Romualdo Formation, which dates to the Albian stage of the Early Cretaceous Period, about 100 million years ago. The formation is part of the Santana Group and, at the time Irritator was described, was thought to be a member of what was then considered the Santana Formation. The Romualdo Formation is a Lagerstätte (a sedimentary deposit that preserves fossils in excellent condition) consisting of lagoonal limestone concretions embedded in shales, and overlies the Crato Formation. It is well-known for preserving fossils three-dimensionally in calcareous concretions, including many pterosaur fossils. In addition to muscle fibres of pterosaurs and dinosaurs, fish preserving gills, digestive tracts, and hearts have been found there.[45][46] The climate of the formation was tropical and largely corresponded to today's Brazilian climate.[47] The sediment layers are dated to a time when the Southern Atlantic Ocean was opening, forming the series of circum-Atlantic basins of southern Brazil and southwestern Africa, but the northeastern part of Brazil and West Africa were still connected.[1]

This environment was dominated by pterosaurs, including Anhanguera, Araripedactylus, Araripesaurus, Brasileodactylus, Cearadactylus, Coloborhynchus, Santanadactylus, Tapejara, Thalassodromeus, Tupuxuara,[48] Barbosania, Maaradactylus,[49] Tropeognathus, and Unwindia.[50] The known dinosaur fauna besides Irritator was represented by other theropods like the tyrannosauroid Santanaraptor, the compsognathid Mirischia,[51] an indeterminate unenlagiine dromaeosaur,[52] and a megaraptoran.[53] The crocodyliforms Araripesuchus and Caririsuchus,[54] as well as the turtles Brasilemys,[55] Cearachelys,[56] Araripemys, Euraxemys,[57] and Santanachelys, are known from the deposits.[58] There were also clam shrimps, sea urchins, ostracods, and molluscs.[13] An abundance of well-preserved fish fossils records the presence of hybodont sharks, guitarfish, gars, amiids, ophiopsids, oshuniids, pycnodontids, aspidorhynchids, cladocyclids, bonefishes, chanids, mawsoniids, and other uncertain forms.[59] Most of its local flora was xerophytic (adapted to dry environments). The most widespread plants were Cycadales, and the extinct conifer Brachyphyllum.[51] The scant vegetation was incapable of sustaining a large population of herbivorous animals, which might explain why the dinosaur fossil record of the region consists mostly if not entirely of predatory theropods. Naish and colleagues suggested that following storm events, pterosaur and fish carcasses might have washed up on the shoreline, and consequently provided theropods with plenty of carrion.[51] Similarities between the fauna of the Romualdo and Crato Formations to transoceanic assemblages in Africa suggest the connection of the Araripe Basin to the Tethys Sea, though this link was likely sporadic, because the lack of marine invertebrates indicates the basin had a non-marine depositional setting.[59]

Many spinosaurs likely had semiaquatic habits, as has been indicated by use of techniques like isotope analysis, where it has been found that they likely took advantage of aquatic prey and environments to occupy a separate ecological niche from other theropods, therefore avoiding competition.[60][61] Spinosaurines in particular appear to have been more adapted for such lifestyles than baryonychines.[53] In May 2018, Aureliano and colleagues conducted an analysis on the Romualdo Formation tibia fragment. CT scanning of the specimen in the University of San Carlos revealed the presence of osteosclerosis (high bone density).[53] This condition had previously only been observed in Spinosaurus, as a possible way of facilitating submersion in water by making its bones heavier.[62] The presence of this condition on the Brazilian leg fragment showed that compact-boned spinosaurids were already present at least 10 million years before the appearance of Spinosaurus aegyptiacus. According to phylogenetic bracketing, osteosclerosis might have been therefore been the norm in spinosaurines.[53] However, the significance of these traits was questioned in an August 2018 publication, where paleobiologist Donald Henderson argued that osteosclerosis would not have changed a theropod's buoyancy to a significant extent.[63]
Taphonomy
The taphonomy (changes between death and fossilization) of Irritator's holotype specimen have been discussed by some researchers. The skull was found lying on its side. Preceding fossilization, several bones from the back of the braincase, as well as the dentary, splenial, coronoid, and right angular bones from the lower jaw were lost. Other bones, mostly from the skull rear, had become disarticulated and displaced towards alternate regions of the head before burial.[64] Naish and colleagues in 2004 asserted that the Romualdo Formation dinosaur fauna is represented by animals that died near shorelines or rivers before being carried out to sea, where their floating remains were eventually fossilized.[42] In 2018, Aureliano and colleagues argued against this scenario, stating that the Irritator holotype's mandible was preserved in articulation with the rest of the skull, whereas it would have likely detached had the carcass been floating at sea. They also noted that the corpse would have quickly sunk due to the osteosclerosis of the skeleton.[53]
References
- 1 2 3 4 5 6 7 8 9 10 11 12 13 Martill, D. M.; Cruickshank, A. R. I.; Frey, E.; Small, P. G.; Clarke, M. (1996). "A new crested maniraptoran dinosaur from the Santana Formation (Lower Cretaceous) of Brazil". Journal of the Geological Society. 153 (1): 5–8. Bibcode:1996JGSoc.153....5M. doi:10.1144/gsjgs.153.1.0005.
- ↑ Gibney, Elizabeth (2014-03-04). "Brazil clamps down on illegal fossil trade". Nature. 507 (7490): 20. doi:10.1038/507020a. ISSN 0028-0836. PMID 24598620.
- 1 2 Machado, E. B., and A. W. A. Kellner. "Notas Sobre Spinosauridae (Theropoda, Dinosauria)", Anuário do Instituto de Geociências - UFRJ, Vol. 28-1, 2005, pp. 158-173.
- ↑ Susanna., Davidson, (2005). The Usborne Internet-linked World atlas of dinosaurs. Turnbull, Stephanie., Firth, Rachel., Rey, Luis V., Slane, Andrea., Naish, Darren., Martill, David M. New York: Scholastic Inc. ISBN 978-0439818407. OCLC 62384519.
- ↑ Malam, John (2007). Encyclopedia of dinosaurs. Parker, Steve, 1952-. Bath, UK: Parragon Pub. ISBN 9781405499231. OCLC 227315325.
- ↑ Don., Lessem, (2004). Scholastic dinosaurs A to Z : the ultimate dinosaur encyclopedia. Sovak, Jan,, Scholastic Reference (Firm). New York: Scholastic. ISBN 978-0439678667. OCLC 63835422.
- ↑ Dougal., Dixon, (2009). The ultimate guide to dinosaurs (North American ed.). Tunbridge Wells, Kent, UK: Ticktock. ISBN 9781846969881. OCLC 428131451.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 Sues, H. D.; Frey, E.; Martill, D. M.; Scott, D. M. (2002). "Irritator challengeri, a spinosaurid (Dinosauria: Theropoda) from the Lower Cretaceous of Brazil". Journal of Vertebrate Paleontology. 22 (3): 535–547. doi:10.1671/0272-4634(2002)022[0535:ICASDT]2.0.CO;2.
- ↑ Isaak, Mark (2008) "Curiosities of Biological Nomenclature". Retrieved 2009-07-26.
- 1 2 3 4 5 6 Sales, Marcos A. F.; Schultz, Cesar L. (2017-11-06). "Spinosaur taxonomy and evolution of craniodental features: Evidence from Brazil". PLOS ONE. 12 (11). doi:10.1371/journal.pone.0187070#pone.0187070.ref015 (inactive 2018-10-07). ISSN 1932-6203.
- 1 2 Carrano, Matthew T.; Benson, Roger B. J.; Sampson, Scott D. (2012-05-17). "The phylogeny of Tetanurae (Dinosauria: Theropoda)". Journal of Systematic Palaeontology. 10 (2): 211–300. doi:10.1080/14772019.2011.630927. ISSN 1477-2019.
- 1 2 3 4 Sasso, Cristiano Dal; Maganuco, Simone; Buffetaut, Eric; Mendez, Marco A. (2005-12-30). "New information on the skull of the enigmatic theropodSpinosaurus, with remarks on its size and affinities". Journal of Vertebrate Paleontology (Submitted manuscript). 25 (4): 888–896. doi:10.1671/0272-4634(2005)025[0888:niotso]2.0.co;2. ISSN 0272-4634.
- 1 2 Mabesoone, J.M.; Tinoco, I.M. (1973-10-01). "Palaeoecology of the Aptian Santana Formation (Northeastern Brazil)". Palaeogeography, Palaeoclimatology, Palaeoecology. 14 (2): 97–118. doi:10.1016/0031-0182(73)90006-0. ISSN 0031-0182.
- ↑ Kellner, Alexander W. A.; Azevedo, Sergio A. K.; Machado, Elaine B.; Carvalho, Luciana B.; Henriques, Deise D. R. (2011). "A new dinosaur (Theropoda, Spinosauridae) from the Cretaceous (Cenomanian) Alcântara Formation, Cajual Island, Brazil" (PDF). Anais da Academia Brasileira de Ciências. 83 (1): 99–108. doi:10.1590/S0001-37652011000100006. ISSN 0001-3765.
- 1 2 3 4 5 6 7 8 Sales, M.A.F.; Schultz, C.L. (2017). "Spinosaur taxonomy and evolution of craniodental features: Evidence from Brazil". PLoS ONE. 12 (11): e0187070. Bibcode:2017PLoSO..1287070S. doi:10.1371/journal.pone.0187070. PMC 5673194. PMID 29107966.
- 1 2 3 4 5 6 7 Aureliano, Tito; Ghilardi, Aline M.; Buck, Pedro V.; Fabbri, Matteo; Samathi, Adun; Delcourt, Rafael; Fernandes, Marcelo A.; Sander, Martin (3 May 2018). "Semi-aquatic adaptations in a spinosaur from the Lower Cretaceous of Brazil". Cretaceous Research (Submitted manuscript). 90: 283–295. doi:10.1016/j.cretres.2018.04.024. ISSN 0195-6671.
- 1 2 Bittencourt, Jonathas; Kellner, Alexander (2004-01-01). "On a sequence of sacrocaudal theropod dinosaur vertebrae from the Lower Cretaceous Santana Formation, northeastern Brazil". Arq Mus Nac. 62: 309–320.
- ↑ Martill, David (1994). "Fake fossils from Brazil". Geology Today. 10 (3): 111–115. doi:10.1111/j.1365-2451.1994.tb00882.x. ISSN 0266-6979.
- 1 2 3 4 Kellner, A.W.A.; Campos, D.A. (1996). "First Early Cretaceous dinosaur from Brazil with comments on Spinosauridae". N. Jb. Geol. Paläont. Abh. 199 (2): 151–166.
- 1 2 (in Portuguese) O Estado de S. Paulo, May 14, 2009, available at ; O Globo, May 15, 2009, abridgement available at ; university's announcement at "Archived copy". Archived from the original on 2011-07-06. Retrieved 2010-01-13.
- 1 2 3 Naish, Darren. "Brilliant Brazilian spinosaurids". Scientific American Blog Network. Retrieved 2018-09-30.
- ↑ Belam, Martin (September 2018). "Brazil's national museum: what could be lost in the fire?". The Guardian. Retrieved 2018-10-07.
- ↑ Naish, Darren. "In Rio for the 2013 International Symposium on Pterosaurs". Scientific American Blog Network. Retrieved 2018-10-07.
- ↑ Charig, A. J.; Milner, A. C. (1997). "Baryonyx walkeri, a fish-eating dinosaur from the Wealden of Surrey". Bulletin of the Natural History Museum of London. 53: 11–70.
- ↑ Sereno, P. C.; Beck, A. L.; Dutheuil, D. B.; Gado, B.; Larsson, H. C.; Lyon, G. H.; Marcot, J. D.; Rauhut, O. W. M.; Sadleir, R. W.; Sidor, C. A.; Varricchio, D. J.; Wilson, G. P.; Wilson, J. A. (1998). "A long-snouted predatory dinosaur from Africa and the evolution of spinosaurids". Science. 282 (5392): 1298–1302. Bibcode:1998Sci...282.1298S. doi:10.1126/science.282.5392.1298. PMID 9812890.
- ↑ Buffetaut, E.; Ouaja, M. (2002). "A new specimen of Spinosaurus (Dinosauria, Theropoda) from the Lower Cretaceous of Tunisia, with remarks on the evolutionary history of the Spinosauridae" (PDF). Bulletin de la Société Géologique de France. 173 (5): 415–421. doi:10.2113/173.5.415.
- ↑ Kellner, A. W. A.; Campos, D. A. (2000). "Brief review of dinosaur studies and perspectives in Brazil". Anais da Academia Brasileira de Ciências. 72 (4): 509–538. doi:10.1590/S0001-37652000000400005.
- ↑ Paul, G.S., 2010, The Princeton Field Guide to Dinosaurs, Princeton University Press p. 87
- ↑ Holtz, Thomas R. Jr. (2011) Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages, Winter 2010 Appendix.
- ↑ Holtz, T.R. Jr. (2014). "Supplementary Information to Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages". University of Maryland. Retrieved 2014-09-05.
- ↑ Dixon, Dougal (2009). The Ultimate Guide to Dinosaurs. Ticktock Books. ISBN 9781846969881.
- ↑ Sales, Marcos (2017). "Contribuições à paleontologia de Terópodes não-avianos do Mesocretáceo do Nordeste do Brasil". 1. Porto Alegre: Universidade Federal do Rio Grande do Sul: 54.
- 1 2 3 4 5 HONE, David William Elliott; HOLTZ, Thomas Richard (2017). "A Century of Spinosaurs - A Review and Revision of the Spinosauridae with Comments on Their Ecology". Acta Geologica Sinica - English Edition. 91 (3): 1120–1132. doi:10.1111/1755-6724.13328. ISSN 1000-9515.
- 1 2 Rayfield, Emily J.; Milner, Angela C.; Xuan, Viet Bui; Young, Philippe G. (2007-12-12). "Functional morphology of spinosaur 'crocodile-mimic' dinosaurs". Journal of Vertebrate Paleontology (Submitted manuscript). 27 (4): 892–901. doi:10.1671/0272-4634(2007)27[892:fmoscd]2.0.co;2. ISSN 0272-4634.
- ↑ Zelenitsky, D. K.; Therrien, F.; Erickson, G. M.; Debuhr, C. L.; Kobayashi, Y.; Eberth, D. A.; Hadfield, F. (2012). "Feathered Non-Avian Dinosaurs from North America Provide Insight into Wing Origins". Science. 338 (6106): 510–514. Bibcode:2012Sci...338..510Z. doi:10.1126/science.1225376. PMID 23112330.
- ↑ Rauhut, Oliver W. M. (2003). The interrelationships and evolution of basal theropod dinosaurs. London, [England]: The Palaeontological Association. ISBN 978-0-901702-79-1.
- ↑ Holtz, Thomas; E. Molnar, Ralph; Currie, Philip (2004-06-12), "Basal Tetanurae", The Dinosauria: Second Edition, pp. 71–110, retrieved 2018-09-25
- ↑ Sereno, P. C.; Beck, A. L.; Dutheuil, D. B.; Gado, B.; Larsson, H. C.; Lyon, G. H.; Marcot, J. D.; Rauhut, O. W. M.; Sadleir, R. W.; Sidor, C. A.; Varricchio, D. J.; Wilson, G. P.; Wilson, J. A. (1998). "A long-snouted predatory dinosaur from Africa and the evolution of spinosaurids". Science. 282 (5392): 1298–1302. Bibcode:1998Sci...282.1298S. doi:10.1126/science.282.5392.1298. PMID 9812890.
- ↑ Carrano, Matthew T.; Benson, Roger B. J.; Sampson, Scott D. (2012-05-17). "The phylogeny of Tetanurae (Dinosauria: Theropoda)". Journal of Systematic Palaeontology. 10 (2): 211–300. doi:10.1080/14772019.2011.630927. ISSN 1477-2019.
- ↑ Benson, R. B. J.; Carrano, M. T.; Brusatte, S. L. (2009). "A new clade of archaic large-bodied predatory dinosaurs (Theropoda: Allosauroidea) that survived to the latest Mesozoic". Naturwissenschaften. 97 (1): 71–78. Bibcode:2010NW.....97...71B. doi:10.1007/s00114-009-0614-x. PMID 19826771.
- ↑ Evers, Serjoscha W.; Rauhut, Oliver W.M.; Milner, Angela C.; McFeeters, Bradley; Allain, Ronan (2015). "A reappraisal of the morphology and systematic position of the theropod dinosaurSigilmassasaurusfrom the "middle" Cretaceous of Morocco". PeerJ. 3: e1323. doi:10.7717/peerj.1323. ISSN 2167-8359. PMC 4614847. PMID 26500829.
- 1 2 Naish, D.; Martill, D. M.; Frey, E. (2004). "Ecology, Systematics and Biogeographical Relationships of Dinosaurs, Including a New Theropod, from the Santana Formation (?Albian, Early Cretaceous) of Brazil". Historical Biology. 16 (2–4): 57–70. CiteSeerX 10.1.1.394.9219. doi:10.1080/08912960410001674200.
- ↑ Buffetaut, E.; Martill, D.; Escuillié3, F. (2004). "Pterosaurs as part of a spinosaur diet". Nature. 429 (33): 33. Bibcode:2004Natur.429...33B. doi:10.1038/430033a. PMID 15229562.
- ↑ Cuff, Andrew R.; Rayfield, Emily J. (2013-05-28). "Feeding Mechanics in Spinosaurid Theropods and Extant Crocodilians". PLoS ONE. 8 (5): e65295. doi:10.1371/journal.pone.0065295. ISSN 1932-6203. PMC 3665537. PMID 23724135.
- ↑ Kellner, A. W. A.; Campos, D. A. (2002). "The function of the cranial crest and jaws of a unique pterosaur from the early Cretaceous of Brazil". Science. 297 (5580): 389–392. doi:10.1126/science.1073186. PMID 12130783.
- ↑ Pêgas, R. V.; Costa, F. R.; Kellner, A. W. A. (2018). "New Information on the osteology and a taxonomic revision of The genus Thalassodromeus (Pterodactyloidea, Tapejaridae, Thalassodrominae)". Journal of Vertebrate Paleontology. 38 (2): e1443273. doi:10.1080/02724634.2018.1443273.
- ↑ Mabesoone, J.M; Tinoco, I.M (1973). "Palaeoecology of the Aptian Santana Formation (Northeastern Brazil)". Palaeogeography, Palaeoclimatology, Palaeoecology. 14 (2): 97–118. doi:10.1016/0031-0182(73)90006-0. ISSN 0031-0182.
- ↑ Barrett, Paul; Butler, Richard; Edwards, Nicholas; R. Milner, Andrew (2008-12-31). "Pterosaur distribution in time and space: An atlas". Zitteliana Reihe B: Abhandlungen der Bayerischen Staatssammlung Fur Palaontologie und Geologie. 28: 61–107.
- ↑ Renan A. M. Bantim; Antônio A. F. Saraiva; Gustavo R. Oliveira; Juliana M. Sayão (2014). "A new toothed pterosaur (Pterodactyloidea: Anhangueridae) from the Early Cretaceous Romualdo Formation, NE Brazil". Zootaxa. 3869 (3): 201–223. doi:10.11646/zootaxa.3869.3.1. PMID 25283914.
- ↑ David M. Martill (2011). "A new pterodactyloid pterosaur from the Santana Formation (Cretaceous) of Brazil". Cretaceous Research. 32 (2): 236–243. doi:10.1016/j.cretres.2010.12.008.
- 1 2 3 Martill, David; Frey, Eberhard; Sues, Hans-Dieter; RI Cruickshank, Arthur (2011-02-09). "Skeletal remains of a small theropod dinosaur with associated soft structures from the Lower Cretaceous Santana Formation of NE Brazil". Canadian Journal of Earth Sciences. 37 (6): 891–900. doi:10.1139/cjes-37-6-891.
- ↑ Candeiro, Carlos Roberto A.; Cau, Andrea; Fanti, Federico; Nava, Willian R.; Novas, Fernando E. (2012-10-01). "First evidence of an unenlagiid (Dinosauria, Theropoda, Maniraptora) from the Bauru Group, Brazil". Cretaceous Research. 37: 223–226. doi:10.1016/j.cretres.2012.04.001. ISSN 0195-6671.
- 1 2 3 4 5 Aureliano, Tito; Ghilardi, Aline M.; Buck, Pedro V.; Fabbri, Matteo; Samathi, Adun; Delcourt, Rafael; Fernandes, Marcelo A.; Sander, Martin (3 May 2018). "Semi-aquatic adaptations in a spinosaur from the Lower Cretaceous of Brazil". Cretaceous Research (Submitted manuscript). 90: 283–295. doi:10.1016/j.cretres.2018.04.024. ISSN 0195-6671.
- ↑ Figueiredo, R. G.; Kellner, A. W. A. (2009). "A new crocodylomorph specimen from the Araripe Basin (Crato Member, Santana Formation), northeastern Brazil". Paläontologische Zeitschrift. 83 (2): 323–331. doi:10.1007/s12542-009-0016-6.
- ↑ de Lapparent de Broin, F. (2000). "The oldest pre-Podocnemidid turtle (Chelonii, Pleurodira), from the early Cretaceous, Ceará state, Brasil, and its environment". Treballs del Museu de Geologia de Barcelona. 9: 43–95. ISSN 2385-4499.
- ↑ Gaffney, Eugene S.; Diogenes de Almeida Campos; Ren Hirayama (2001-02-27). "Cearachelys, a New Side-necked Turtle (Pelomedusoides: Bothremydidae) from the Early Cretaceous of Brazil". American Museum Novitates. 3319: 1–20. doi:10.1206/0003-0082(2001)319<0001:CANSNT>2.0.CO;2. hdl:2246/2936.
- ↑ S GAFFNEY, EUGENE; Tong, Haiyan; A MEYLAN, PETER (2009-09-02). "Evolution of the side-necked turtles: The families Bothremydidae, Euraxemydidae, and Araripemydidae". Bulletin of the American Museum of Natural History. 300: 1–698. doi:10.1206/0003-0090(2006)300[1:EOTSTT]2.0.CO;2. hdl:2246/5824.
- ↑ Hirayama, Ren (1998). "Oldest known sea turtle". Nature. 392 (6677): 705–708. doi:10.1038/33669. ISSN 0028-0836.
- 1 2 Brito, Paulo; Yabumoto, Yoshitaka (2011). "An updated review of the fish faunas from the Crato and Santana formations in Brazil, a close relationship to the Tethys fauna". Bulletin of Kitakyushu Museum of Natural History and Human History, Ser. A. 9.
- ↑ Amiot, R.; Buffetaut, E.; Lécuyer, C.; Wang, X.; Boudad, L.; Ding, Z.; Fourel, F.; Hutt, S.; Martineau, F.; Medeiros, A.; Mo, J.; Simon, L.; Suteethorn, V.; Sweetman, S.; Tong, H.; Zhang, F.; Zhou, Z. (2010). "Oxygen isotope evidence for semi-aquatic habits among spinosaurid theropods". Geology. 38 (2): 139–142. Bibcode:2010Geo....38..139A. doi:10.1130/G30402.1.
- ↑ Hassler, A.; Martin, J. E.; Amiot, R.; Tacail, T.; Godet, F. Arnaud; Allain, R.; Balter, V. (2018-04-11). "Calcium isotopes offer clues on resource partitioning among Cretaceous predatory dinosaurs". Proc. R. Soc. B. 285 (1876): 20180197. doi:10.1098/rspb.2018.0197. ISSN 0962-8452. PMC 5904318. PMID 29643213.
- ↑ Ibrahim, N.; Sereno, P. C.; Dal Sasso, C.; Maganuco, S.; Fabbri, M.; Martill, D. M.; Zouhri, S.; Myhrvold, N.; Iurino, D. A. (2014). "Semiaquatic adaptations in a giant predatory dinosaur". Science. 345 (6204): 1613–1616. doi:10.1126/science.1258750. PMID 25213375.
- ↑ Henderson, Donald M. (2018-08-16). "A buoyancy, balance and stability challenge to the hypothesis of a semi-aquatic Spinosaurus Stromer, 1915 (Dinosauria: Theropoda)". PeerJ. 6: e5409. doi:10.7717/peerj.5409. ISSN 2167-8359. PMC 6098948. PMID 30128195.
- ↑ Sues, H. D.; Frey, E.; Martill, D. M.; Scott, D. M. (2002). "Irritator challengeri, a spinosaurid (Dinosauria: Theropoda) from the Lower Cretaceous of Brazil". Journal of Vertebrate Paleontology. 22 (3): 535–547. doi:10.1671/0272-4634(2002)022[0535:ICASDT]2.0.CO;2.
Further reading
- Don Lessem (2003). Dinosaurs A to Z. Scholastic New York. ISBN 978-0-439-16591-4. .
External links
| Wikimedia Commons has media related to Irritator. |
- Irritator at The Theropod Database
- Diagram of the skull pieces found from Irritator
- Angaturama(Irritator) on post stamp
| |||||||||||||||||||||
| Piatnitzkysauridae | 
.png) | ||||||||||||||||||||
| Megalosauria |
| ||||||||||||||||||||
.jpg)