Haplogroup N-M231
| Haplogroup N | |
|---|---|
.PNG) | |
| Possible time of origin |
36,800 [95% CI 34,300–39,300] years before present (YFull[1]) 44,700 or 38,300 ybp depending on mutation rate[2] 41,900 [95% CI 40,175-43,591] ybp[3] |
| Coalescence age |
22,100 [95% CI 20,000–24,400] ybp (YFull[1]) 25,313 [95% CI 21,722-28,956] ybp[3] |
| Possible place of origin | East Asia[4][5] |
| Ancestor | NO |
| Defining mutations | M231 |
| Highest frequencies | Nganassan 92%, Yakuts 75%, Nenets 75%, Finns 60%, Baltic States 45% (McDonald 2005), Saami 40% |
Haplogroup N (M231) is a Y-chromosome DNA haplogroup defined by the presence of the single-nucleotide polymorphism (SNP) marker M231.[Phylogenetics 1]
It is most commonly found in males originating from northern Eurasia. It also has been observed at lower frequencies in populations native to other regions, including the Balkans, East Asia, Central Asia and the Pacific.
Origins
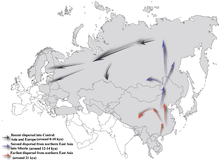
Haplogroup NO-M214 – its most recent common ancestor with its sibling, haplogroup O-M175 – is estimated to have existed about 36,800–44,700 years ago.[1][7][2]
It is generally considered that N-M231 arose in South-East Asia approximately 19,400 (±4,800) years ago and re-populated northern Eurasia after the Last Glacial Maximum. Males carrying the marker apparently moved northwards as the climate warmed in the Holocene, migrating in a counter-clockwise path (through modern China and Mongolia), to eventually become concentrated in areas as far away as Fennoscandia and the Baltic.(Rootsi 2006). The apparent dearth of haplogroup N-M231 amongst Native American peoples indicates that it spread after Beringia was submerged (Chiaroni 2009), about 11,000 years ago.
Distribution

Haplogroup N has a wide geographic distribution throughout northern Eurasia, and it also has been observed occasionally in other areas, including South-East Asia, the Pacific, Central Asia and the Balkans.
Its highest frequency occurs among ethnic Finnic peoples and Baltic-speaking peoples of northern Europe, the Ob-Ugric-speaking and Northern Samoyed peoples of western Siberia, and the Siberian Turkic-speaking Yakuts (McDonald 2005). Nearly all members of haplogroup N among these populations of northern Eurasia belong to subclades of N1a-F1206/M2013/S11466.
Y-chromosomes belonging to N1b-F2930/M1881/V3743, or N1*-CTS11499/L735/M2291(xN1a-F1206/M2013/S11466), have been found at relatively high levels in South China and adjoining areas of southeastern Asia.
N2-Y6503, the other primary subclade of haplogroup N, is extremely rare and is mainly represented among extant humans by a recently formed subclade that is virtually restricted to the countries making up the former Yugoslavia (Bosnia-Herzegovina, Croatia, Serbia, and Montenegro). Other members of N2-Y6503 include a Hungarian with recent ancestry from Suceava in Bukovina, a Slovakian, a few British individuals, and an Altaian.[1]
N* (M231)
Y-chromosomes that display the M231 mutation that defines Haplogroup N-M231, but do not display the CTS11499, L735, M2291 mutations that define Haplogroup N1 are said to belong to paragroup N-M231*.[4]
N-M231* has been found at low levels in China and Cambodia.[4] Out of a sample of 165 Han males from China, two individuals (1.2%) were found to belong to N*.(Karafet 2010).[Footnote 1] One originated from Guangdong and one from Shaanxi.
N1 (CTS11499; Z4762; CTS3750)
In 2014, CTS11499/L735/M2291 replaced LLY22g was retired as a defining SNP for Haplogroup N1; it was replaced by . According to ISOGG, LLY22g is problematic because it is a "palindromic marker and can easily be misinterpreted".[4] Consequently, the position of many previously examples of "N-LLY22g", within N-M231 has become unclear.
N1* has been reported to reach a frequency of up to 30% (13/43) among the Yi people of Butuo County, Sichuan in Southwest China (Hammer 2005, Karafet 2001, and Wen2004b). It is also found in 34.6% of Lhoba people (Wen 2004, Bo Wen 2004).[8]
Paragroup N-LLY22g* also has been found in samples of Han Chinese, but with widely varying frequency:
- 15.0% (6/40) Guangdong Han (Hammer 2005 and Karafet 2001)
- 6.8% (3/44) Shaanxi Han (Hammer 2005 and Karafet 2001)
- 6.7% (2/30) Han from Lanzhou (Xue 2006)
- 3.6% (3/84) Taiwanese Han (Hammer 2005)
- 2.9% (1/34) Han from Chengdu (Xue 2006)
- 2.9% (1/35) Han from Harbin (Xue 2006)
- 2.9% (1/35) Han from Meixian District (Xue 2006)
- 0% (0/32) Han from Yining City (Xue 2006)
Other populations in which representatives of N1 * have been found include:
- Hani people (4/34 = 11.8%) (Xue 2006)
- Sibe people (4/41 = 9.8%) (Xue 2006)
- Tujia people (2/49 = 4.1%) (Hammer 2005)
- Manchu people (2/52 = 3.8% (Hammer 2005) to 2/35 = 5.7% (Xue 2006)
- Bit people (1/28 = 3.6%) (Cai 2011)
- Uyghurs (2/70 = 2.9% (Xue 2006) to 2/67 = 3.0%) (Hammer 2005)
- Tibetan people (3/105 = 2.9% (Hammer 2005) to 3/35 = 8.6% (Xue 2006))
- Koreans (0/106 = 0.0% – 2/25 = 8% (Rootsi 2006, Xue 2006, and Kim 2007)
- Vietnamese people (2/70 = 2.9%) (Hammer 2005)
- Japanese people (0/70 Tokushima – 2/26 = 7.7% Aomori) (Hammer 2005)
- Manchurian Evenks (0/26 = 0.0% (Xue 2006) to 1/41 = 2.4%(Hammer 2005))
- Altai people (0/50 Northern to 5/96 = 5.2% Southern, or 0/43 Beshpeltir to 5/46 = 10.9% Kulada),(Hammer 2005)(Kharkov 2007)
- Shors (2/23 = 8.7%) (Rootsi 2006)
- Khakas people (5/181 = 2.8%) (Rootsi 2006)
- Tuvans (5/311 = 1.6%) (Rootsi 2006)
- Southern Borneo (1/40 = 2.5%) (Rootsi 2006)
- Forest Nenets (1/89 = 1.1%) (Rootsi 2006)
- Yakuts (0/215 – 1/121 = 0.8%) (Rootsi 2006)
- Turkish people (1/523 = 0.2%) (Rootsi 2006) In Turkey, the total of subclades of haplogroup N-M231 amounts to 4% of the male population.
- One individual who belongs either to N* or N1* has been found in a sample of 77 males from Kathmandu, Nepal (1/77 = 1.3% N-M231(xM128,P43,Tat)) (Gayden 2007).
N1(xN1a, N1c) was found in ancient bones of Liao civilization:[9]
- Niuheliang (Hongshan Culture, 6500–5000 BP) 66.7%(=4/6)
- Halahaigou (Xiaoheyan Culture, 5000–4200 BP) 100.0%(=12/12)
- Dadianzi (Lower Xiajiadian culture, 4200–3600 BP) 60.0%(=3/5)
N1a (F1206/M2013/S11466)
The N1a2-F1008/L666 clade and N1a1-M46/Page70/Tat are estimated to share a most recent common ancestor in N1a-F1206/M2013/S11466 approximately 15,900 [95% CI 13,900 <-> 17,900] years before present[1] or 17,621 [95% CI 14,952 <-> 20,282] years before present.[3]
N1a1 (M46/Page70/Tat, L395/M2080)
The mutations that define the subclade N-M46[Phylogenetics 2] are M46/Tat and P105. This is the most frequent subclade of N. It arose probably in the region of present-day China, and subsequently experienced serial bottlenecks in Siberia and secondary expansions in eastern Europe (Rootsi 2006). Haplogroup N-M46 is approximately 14,000 years old.
In Siberia, haplogroup N-M46 reaches a maximum frequency of approximately 90% among the Yakuts, a Turkic people who live mainly in the Sakha (Yakutia) Republic. However, N-M46 is absent or present with much lower frequency among many of the Yakuts' neighbors, such as Evenks and Evens.[10] It also has been detected in 5.9% (3/51) of a sample of Hmong Daw from Laos (Cai 2011), 2.4% (2/85) of a sample from Seoul, South Korea (Katoh 2004), and in 1.4% (1/70) of a sample from Tokushima, Japan (Hammer 2005).
The haplogroup N-M46 has a low diversity among Yakuts suggestive of a population bottleneck or founder effect ( & Pakendorf 2002). This was confirmed by a study of ancient DNA which traced the origins of the male Yakut lineages to a small group of horse-riders from the Cis-Baikal area (Crubézy 2010).
N1a1a (M178)
The subclade N-M178[Phylogenetics 3] is defined by the presence of markers M178 and P298. N-M178* has higher average frequency in Northern Europe than in Siberia, reaching frequencies of approximately 60% among Finns and approximately 40% among Latvians, Lithuanians & 35% among Estonians (Derenko 2007 and Lappalainen 2008).
Miroslava Derenko and her colleagues noted that there are two subclusters within this haplogroup, both present in Siberia and Northern Europe, with different histories. The one that they labelled N3a1 first expanded in south Siberia (approximately 10,000 years ago on their calculated by the Zhivotovsky method) and spread into Northern Europe where its age they calculated as around 8,000 years ago. Meanwhile, the younger subcluster, which they labelled N3a2, originated in south Siberia (probably in the Baikal region) approximately 4,000 years ago (Derenko 2007).
N1a2 (F1008/L666)
N1a2a-M128 and N1a2b-B523/P43 are estimated to share a most recent common ancestor in N1a2-F1008/L666 approximately 8,800 [95% CI 7,700 <-> 10,000] years before present[1] or 9,314 [95% CI 7,419 <-> 11,264] years before present.[3]
At least three of six tested male specimens from the "Early Neolithic" (ceramic-using hunter-gatherer of approximately 7200–6200 years before present) layer at the Shamanka archaeological site near the southern end of Lake Baikal have been found to belong to N1a2-L666.[11]
N1a2a-M128
| Haplogroup N-M128 | |
|---|---|
| Possible place of origin | Asia |
| Ancestor | N1c2 (F1008/L666) |
| Defining mutations | M128 |
This subclade is defined by the presence of the marker M128.[Phylogenetics 4] N-M128 was first identified in a sample from Japan (1/23 = 4.3%) and in a sample from Central Asia and Siberia (1/184 = 0.5%) in a preliminary survey of worldwide Y-DNA variation.[12] Subsequently, it has been found with low frequency in some samples of the Manchu people, Sibe people, Evenks, Koreans, Han Chinese, Hui, Tibetans, Vietnamese, Bouyei people, Kazakhs, Uzbeks, the Buzava tribe of Kalmyks,[13] Khakas, and Komis.[14]
A number of Han Chinese, an Ooled Mongol, a Qiang, and a Tibetan were found to belong to a sister branch (or branches) of N-M128 under paragroup N-F1154*.[15]
N1a2b (P43)
Haplogroup N-P43[Phylogenetics 5] is defined by the presence of the marker P43. It has been estimated to be about five thousand years old (TMRCA 4,700 [95% CI 3,800 <-> 5,600] ybp[1] or 4,727 [95% CI 3,824 <-> 5,693] years before present[3]). It is found frequently among Northern Samoyedic peoples; also found at low to moderate frequency among some other Uralic peoples, Turkic peoples, Mongolic peoples, Tungusic peoples, and Siberian Yupik people.
The highest frequencies of N-P43 are observed among north-west Siberian populations: 92% (35/38)[16] in a sample of Nganasan, 78% (7/9)[17][18] in a sample of Enets, 78% (21/27)[19] in a sample of Khants, 75% (44/59)[16] in a sample of Tundra Nenets, 69% (29/42)[3] in another sample of Nenets, 60% (15/25)[20] in a sample of Mansi, 57% (64/112)[21] in another sample of Khants, 54% (27/50)[3] in another sample of Nganasan, 45% (40/89)[16] in a sample of Forest Nenets, 38% (18/47)[22] in a third sample of Khants, and 25% (7/28)[20] in a fourth sample of Khants. In Europe, the N-P43 types have their highest frequency of 20% among Volga-Uralic populations. The extreme western border of the spread of N-P43 is Finland, where this haplogroup occurs only at marginal frequency – 0.4%. Yet N-P43 is quite frequent among Vepsas (17.9%), a small Finnic population living in immediate proximity to Finns, Karelians and Estonians.[23]
Haplogroup N-P43 forms two distinctive subclusters of STR haplotypes, Asian and European, the latter mostly distributed among Uralic-speaking peoples and related populations (Rootsi 2006).
N1a2b1-B478
The TMRCA of N-B478 has been estimated to be 3,007 [95% CI 2,171 <-> 3,970] years before present.[3] It is one of the most prevalent Y-DNA haplogroups among indigenous populations of northwestern Siberia: 69.0% (29/42) Nenets, 50.0% (25/50) Nganasan, 22.2% (12/54) Dolgan from Taymyr, 7.0% (3/43) Selkup, 1.6% (1/63) Ob-Ugrian. It is also quite prevalent among populations of Central Siberia, Southern Siberia, and Mongolia: 17.9% (17/95) Tuvan, 15.5% (27/174) Khakas, 8.7% (2/23) Shor, 8.3% (2/24) Even, 8.2% (5/61) Altaian, 5.3% (3/57) Evenk, 5.0% (19/381) Mongol, 4.2% (9/216) Yakut, 0.9% (1/111) Buryat. A geographically outlying member has been found in a sample of Chuvash (1/114 = 0.88%).[3]
N1b (F2930)
Haplogroup N1b has been predominantly found in populations of southwestern China.[24] However, it also has been found in people all over China as well as in Poland, India, Bhutan, Japan, Vietnam, and Cambodia.
N2 (Y6503)
N2 (Y6503/FGC28528; B482/FGC28394/Y6584) – a primary branch of haplogroup N-M231, is found mostly in Serbia, Croatia, and Bosnia and Herzegovina.[25] [26]
Phylogeny
Phylogenetic tree
In the following tree the nomenclature of three sources is separated by slashes: ISOGG Tree 10 December 2017 (ver.12.317)
- NO-M214
- N M231/Page91, M232/M2188
- N1-Z4762/CTS11499/L735/M2291
- N1a-L729
- N1a1-M46/Page70/Tat
- N1a1a-M178
- N1a1a2-Y23747 Japan,[3][1][27] Oroqen,[1][28] Daur,[28] Hebei,[1] Tibet (Shigatse)[29]
- N1a1a1-F1419
- N-Y24317
- N1a1a1a-L708
- N1a1a1a2-B211 Udmurt,[3] Komi,[3] Chuvash,[3] Ob-Ugrians,[3] Mari,[3] Mordva,[3] Altaian,[3] Belarusian,[3] Karanogay,[3] Karelian,[3] Bashkir,[3] Tatar,[27] Russian,[3][27] Khakas[3]
- N1a1a1a1-P298
- N1a1a1a1b-M2118
- N1a1a1a1a-L392
- N1a1a1a1a1-CTS10760
- N1a1a1a1a1c-B479 Nanai[3]
- N1a1a1a1a1b-PH1266/Y28526/F4134
- N1a1a1a1a1a-CTS2929/VL29 Found with high frequency among Lithuanians, Latvians, Estonians, northwestern Russians, Swedish Saami, Karelians, Nenetses, Finns, and Maris, moderate frequency among other Russians, Belarusians, Ukrainians, and Poles, and low frequency among Komis, Mordva, Tatars, Chuvashes, Dolgans, Vepsa, Selkups, Karanogays, and Bashkirs[3]
- N1a1a1a1a1a1-Z4908
- N1a1a1a1a1a1a-L550/S431
- N1a1a1a1a1a1a1-B215/L1025
- N1a1a1a1a1a1a-L550/S431
- N1a1a1a1a1a2-CTS9976
- N1a1a1a1a1a1-Z4908
- N1a1a1a1a2-Z1936,CTS10082 Found with high frequency among Finns, Vepsa, Karelians, Swedish Saami, northwestern Russians, Bashkirs, and Volga Tatars, moderate frequency among other Russians, Komis, Nenetses, Ob-Ugrians, Dolgans, and Siberian Tatars, and low frequency among Mordva, Nganasans, Chuvashes, Estonians, Latvians, Ukrainians, and Karanogays[3]
- N1a1a1a1a2a-Z1928/CTS2733
- N-YP6092
- N-B195
- N1a1a1a1a2a-Z1925
- N-Z1925*
- N-Y29767
- N1a1a1a1a2a2a1a1-Z1926
- N1a1a1a1a2a1c-PH3340/Y13850
- N1a1a1a1a2a1c1-L1034
- N-Y28538
- N-L1442
- N1a1a1a1a2a1c2-Y24361
- N1a1a1a1a2a1c1-L1034
- N-YP6092
- N1a1a1a1a2a-Z1928/CTS2733
- N1a1a1a1a3-B197/Y16323
- N1a1a1a1a3a-F4205 Found with high frequency among Buryats,[3] moderate frequency among Karanogays,[3] Tuvans,[3] and Mongols,[3] and low frequency among Altaians,[3] Siberian Tatars,[3] Kazakhs,[3] Evenks,[3] Crimean Tatars,[3] Karakalpaks,[3] Uzbeks,[3] and Ukrainians[3]
- N1a1a1a1a3b-B202 Found with high frequency among Chukchis,[3] Koryaks,[3] and Siberian Eskimos[3]
- N1a1a1a1a1-CTS10760
- N1a1a-M178
- N1a2-F1008/L666
- N1a2a-M128
- N1a2a1-F710
- N1a2b-B523(P43)
- N1a2b1-B478 (P63) Nenets, Nganasans, Dolgans, Tuvans, Khakasses, Shorians, Evens, Altaians, Selkups, Evenks, Mongols, Yakuts, Ob-Ugrians, Buryats, Chuvashes
- N1a2b1a-B168 Evens
- N1a2b1b-B169
- N1a2b1b1-B170 Nenets, Khanty-Mansi Autonomous Okrug, Inner Mongolia
- N1a2b1b2-B175 Tuvinians, Mongols, Evenks, Yakuts, Tomsk Oblast
- N1a2b2-FGC10872/Y3195
- N1a2b2a-FGC10847/Y3185 (L1419) Vepsas, Maris, Russians (Arkhangelsk Oblast), Komis, Perm Krai, Komi Republic, Ob-Ugrians, Chuvashes, Tatars, Bashkirs, Karelians, Western Finland Province, Tuvans, Buryats, Khakasses, Nganasans, Asian Eskimos
- N1a2b2b-Y23786
- N1a2b2b* Mansis
- N1a2b2b1-B528/Y24384 Udmurts, Komis, Khanties, Tatars, Asian Eskimos, Kirov Oblast, Perm Krai, Medny Island
- N1a2b3-B525 Turkey, Tatars, Bashkirs, Kazakhs, Mongols, Slovakia, Ukrainians, Belarusians, Russians, Afghanistan, Arabs
- N1a2b1-B478 (P63) Nenets, Nganasans, Dolgans, Tuvans, Khakasses, Shorians, Evens, Altaians, Selkups, Evenks, Mongols, Yakuts, Ob-Ugrians, Buryats, Chuvashes
- N1a2a-M128
- N1a1-M46/Page70/Tat
- N1b-F2905
- N1a-L729
- N2-Y6503 Balkans (virtually restricted to the former Yugoslavia)
- N1-Z4762/CTS11499/L735/M2291
- N M231/Page91, M232/M2188
Phylogenetic history
Prior to 2002, there were in academic literature at least seven naming systems for the Y-Chromosome Phylogenetic tree. This led to considerable confusion. In 2002, the major research groups came together and formed the Y-Chromosome Consortium (YCC). They published a joint paper that created a single new tree that all agreed to use. Later, a group of citizen scientists with an interest in population genetics and genetic genealogy formed a working group to create an amateur tree aiming at being above all timely. The table below brings together all of these works at the point of the landmark 2002 YCC Tree. This allows a researcher reviewing older published literature to quickly move between nomenclatures.
| YCC 2002/2008 (Shorthand) | (α) | (β) | (γ) | (δ) | (ε) | (ζ) | (η) | YCC 2002 (Longhand) | YCC 2005 (Longhand) | YCC 2008 (Longhand) | YCC 2010r (Longhand) | ISOGG 2006 | ISOGG 2007 | ISOGG 2008 | ISOGG 2009 | ISOGG 2010 | ISOGG 2011 | ISOGG 2012 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N-LLY22g | 12 | VIII | 1U | 25 | Eu16 | H5 | F | N* | N | N1 | N1 | - | - | - | - | - | - | - |
| N-M128 | 12 | VIII | 1U | 25 | Eu16 | H5 | F | N1 | N1 | N1a | N1a | - | - | - | - | - | - | - |
| N-P63 | 12 | VIII | 1U | 25 | Eu16 | H5 | F | N2 | N2a | N1b1 | N1b1 | - | - | - | - | - | - | - |
| N-TAT | 12 | VIII | 1I | 26 | Eu13 | H5 | F | N3* | N3 | N1c | N1c | - | - | - | - | - | - | - |
| N-M178 | 16 | VIII | 1I | 26 | Eu14 | H5 | F | N3a* | M178 | N1c1 | N1c1 | - | - | - | - | - | - | - |
| N-P21 | 16 | VIII | 1I | 26 | Eu14 | H5 | F | N3a1 | N3a1 | N1c1a | N1c1a | - | - | - | - | - | - | - |
Sources The following research teams per their publications were represented in the creation of the YCC Tree.
Unreliable mutations (SNPs and UEPs)
The b2/b3 deletion in the AZFc region of the Y-chromosome appears to have occurred independently on at least four different occasions. Therefore, this deletion should not be taken as a unique event polymorphism defining this branch of the Y-chromosome tree (ISOGG 2012).
Genetics
- genetic genealogy
- Genetic history of Europe
- Haplogroup
- Haplotype
- Human Y-chromosome DNA haplogroup
- molecular phylogeny
- Paragroup
- Subclade
- Y-chromosome haplogroups in populations of the world
- Y-DNA haplogroups in populations of Europe
- Y-DNA haplogroups in populations of East and Southeast Asia
- Y-DNA haplogroups by ethnic group
Y-DNA N subclades
- N-M231
Y-DNA backbone tree
| Phylogenetic tree of human Y-chromosome DNA haplogroups [χ 1][χ 2] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| "Y-chromosomal Adam" | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A00 | A0-T [χ 3] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A0 | A1 [χ 4] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A1a | A1b | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A1b1 | BT | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| B | CT | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DE | CF | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| D | E | C | F | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| F1 | F2 | F3 | GHIJK | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| G | HIJK | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IJK | H | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IJ | K | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| I | J | LT [χ 5] | K2 [χ 6] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| L | T | K2a [χ 7] | K2b [χ 8] | K2c | K2d | K2e [χ 9] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| K-M2313 [χ 10] | K2b1 [χ 11] | P [χ 12] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| NO | S [χ 13] | M [χ 14] | P1 | P2 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| N | O | Q | R | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
References
Footnotes
- ↑ In Karafet 2010
Work cited
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 YFull Haplogroup YTree v6.05.11 at 25 September 2018.
- 1 2 G. David Poznik, Yali Xue, Fernando L. Mendez, et al. (2016), "Punctuated bursts in human male demography inferred from 1,244 worldwide Y-chromosome sequences." Nature Genetics 2016 June ; 48(6): 593–599. doi:10.1038/ng.3559.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 Ilumäe 2016
- 1 2 3 4 ISOGG, 2016, Y-DNA Haplogroup N and its Subclades – 2016 22 August 2016).
- ↑ (Rootsi 2006)
- 1 2 Shi, H; Qi, X; Zhong, H; Peng, Y; Zhang, X; et al. (2013). "Genetic Evidence of an East Asian Origin and Paleolithic Northward Migration of Y-chromosome Haplogroup N". PLoS ONE. 8 (6): e66102. Bibcode:2013PLoSO...866102S. doi:10.1371/journal.pone.0066102. PMC 3688714. PMID 23840409.
- 1 2 3 Karmin, Monika; Saag, Lauri; Vicente, Mário; et al. (2015). "A recent bottleneck of Y chromosome diversity coincides with a global change in culture". Genome Research. 25 (4): 459–466. doi:10.1101/gr.186684.114.
- ↑ Bo Wen 2004, Analyses of Genetic Structure of Tibeto-Burman Populations Reveals Sex-Biased Admixture in Southern Tibeto-Burmans
- ↑ Yinqiu Cui, Hongjie Li, Chao Ning, Ye Zhang, Lu Chen, Xin Zhao, Erika Hagelberg and Hui Zhou (2013)"Y Chromosome analysis of prehistoric human populations in the West Liao River Valley, Northeast China. " BMC 13:216
- ↑ Duggan, AT; Whitten, M; Wiebe, V; Crawford, M; Butthof, A; et al. (2013). "Investigating the Prehistory of Tungusic Peoples of Siberia and the Amur-Ussuri Region with Complete mtDNA Genome Sequences and Y-chromosomal Markers". PLoS ONE. 8 (12): e83570. Bibcode:2013PLoSO...883570D. doi:10.1371/journal.pone.0083570.
- ↑ Peter de Barros Damgaard, Rui Martiniano, Jack Kamm, et al. (2018), "The first horse herders and the impact of early Bronze Age steppe expansions into Asia." Science 10.1126/science.aar7711 (2018).
- ↑ Peter A. Underhill, Peidong Shen, Alice A. Lin et al., "Y chromosome sequence variation and the history of human populations," Nature Genetics • Volume 26 • November 2000
- ↑ Boris Malyarchuk, Miroslava Derenko, Galina Denisova, Sanj Khoyt, Marcin Woźniak, Tomasz Grzybowski, and Ilya Zakharov, "Y-chromosome diversity in the Kalmyks at the ethnical and tribal levels." Journal of Human Genetics (2013) 58, 804–811; doi:10.1038/jhg.2013.108; published online 17 October 2013.
- ↑ Siiri Rootsi, Lev A Zhivotovsky, Marian Baldovič, et al., "A counter-clockwise northern route of the Y-chromosome haplogroup N from Southeast Asia towards Europe." European Journal of Human Genetics (2007) 15, 204–211. doi:10.1038/sj.ejhg.5201748; published online 6 December 2006.
- ↑ Kang Hu 2015
- 1 2 3 Siiri Rootsi, Lev A Zhivotovsky, Marian Baldovič, et al. (2007), "A counter-clockwise northern route of the Y-chromosome haplogroup N from Southeast Asia towards Europe." European Journal of Human Genetics (2007) 15, 204–211.
- ↑ Tatiana M. Karafet, Ludmila P. Osipova, Marina A. Gubina, Olga L. Posukh, Stephen L. Zegura, and Michael F. Hammer, "High Levels of Y-Chromosome Differentiation among Native Siberian Populations and the Genetic Signature of a Boreal Hunter-Gatherer Way of Life." Human Biology, December 2002, v. 74, no. 6, pp. 761–789.
- ↑ Kristiina Tambets, Bayazit Yunusbayev, Georgi Hudjashov, et al., "Genes reveal traces of common recent demographic history for most of the Uralic-speaking populations." Genome Biology (2018) 19:139. https://doi.org/10.1186/s13059-018-1522-1
- ↑ Sheyla Mirabal, Maria Regueiro, Alicia M Cadenas, L Luca Cavalli-Sforza, Peter A Underhill, Dmitry A Verbenko, Svetlana A Limborska, and Rene J Herrera, "Y-Chromosome distribution within the geo-linguistic landscape of northwestern Russia." European Journal of Human Genetics (2009) 17, 1260–1273; doi:10.1038/ejhg.2009.6.
- 1 2 Ville N Pimenoff, David Comas, Jukka U Palo, Galina Vershubsky, Andrew Kozlov, and Antti Sajantila, "Northwest Siberian Khanty and Mansi in the junction of West and East Eurasian gene pools as revealed by uniparental markers." European Journal of Human Genetics (2008) 16, 1254–1264; doi:10.1038/ejhg.2008.101.
- ↑ KHARKOV, Vladimir Nikolaevich, "СТРУКТУРА И ФИЛОГЕОГРАФИЯ ГЕНОФОНДА КОРЕННОГО НАСЕЛЕНИЯ СИБИРИ ПО МАРКЕРАМ Y-ХРОМОСОМЫ," Genetika 03.02.07 and "АВТОРЕФЕРАТ диссертации на соискание учёной степени доктора биологических наук, Тomsk 2012
- ↑ Kristiina Tambets, Siiri Rootsi, Toomas Kivisild, et al. (2004), "The Western and Eastern Roots of the Saami—the Story of Genetic “Outliers” Told by Mitochondrial DNA and Y Chromosomes." American Journal of Human Genetics 74:661–682.
- ↑ https://www.nature.com/ejhg/journal/v15/n2/full/5201748a.html
- ↑ Hu, et al. 2015. The dichotomy structure of Y chromosome Haplogroup N. arXiv:1504.06463
- ↑ ISOGG, 2018, Y-DNA Haplogroup N and its Subclades - 2018 (24 June 2018).
- ↑ YFull, 2018, N-P189.2 (24 June 2018).
- 1 2 3 4 5 6 7 Haplogroup N North Eurasian YDNA Project at Family Tree DNA
- 1 2 Sebastian Lippold, Hongyang Xu, Albert Ko, Mingkun Li, Gabriel Renaud, Anne Butthof, Roland Schröder, and Mark Stoneking, "Human paternal and maternal demographic histories: insights from high-resolution Y chromosome and mtDNA sequences." Investigative Genetics 2014, 5:13. http://www.investigativegenetics.com/content/5/1/13
- ↑ Wang, LX., Lu, Y., Zhang, C., et al., "Reconstruction of Y-chromosome phylogeny reveals two neolithic expansions of Tibeto-Burman populations." Molecular Genetics and Genomics (2018). https://doi.org/10.1007/s00438-018-1461-2
Journals
- Cai, Xiaoyun; Qin, Zhendong; Wen, Bo; Xu, Shuhua; Wang, Yi; Lu, Yan; Wei, Lanhai; Wang, Chuanchao; et al. (2011). O'Rourke, Dennis, ed. "Human Migration through Bottlenecks from Southeast Asia into East Asia during Last Glacial Maximum Revealed by Y Chromosomes". PLoS ONE. 6 (8): e24282. Bibcode:2011PLoSO...624282C. doi:10.1371/journal.pone.0024282. PMC 3164178. PMID 21904623.
- Chiaroni, Jacques; Underhill, Peter A.; Cavalli-Sforza, Luca L. (2009). "Y chromosome diversity, human expansion, drift, and cultural evolution". Proceedings of the National Academy of Sciences. 106 (48): 20174–79. Bibcode:2009PNAS..10620174C. doi:10.1073/pnas.0910803106. PMC 2787129. PMID 19920170.
- Crubézy, Eric; Amory, Sylvain; Keyser, Christine; Bouakaze, Caroline; Bodner, Martin; Gibert, Morgane; Röck, Alexander; Parson, Walther; Alexeev, Anatoly; Ludes, Bertrand (2010). "Human evolution in Siberia: From frozen bodies to ancient DNA". BMC Evolutionary Biology. 10: 25. doi:10.1186/1471-2148-10-25. PMC 2829035. PMID 20100333.
- Derenko, Miroslava; Malyarchuk, Boris; Denisova, Galina; Wozniak, Marcin; Grzybowski, Tomasz; Dambueva, Irina; Zakharov, Ilia (2007). "Y-chromosome haplogroup N dispersals from south Siberia to Europe". Journal of Human Genetics. 52 (9): 763–70. doi:10.1007/s10038-007-0179-5. PMID 17703276.
- Gayden, Tenzin; Cadenas, Alicia M.; Regueiro, Maria; Singh, Nanda B.; Zhivotovsky, Lev A.; Underhill, Peter A.; Cavalli-Sforza, Luigi L.; Herrera, Rene J. (2007). "The Himalayas as a Directional Barrier to Gene Flow". The American Journal of Human Genetics. 80 (5): 884–94. doi:10.1086/516757. PMC 1852741. PMID 17436243.
- Hammer, Michael F.; Karafet, Tatiana M.; Park, Hwayong; Omoto, Keiichi; Harihara, Shinji; Stoneking, Mark; Horai, Satoshi (2005). "Dual origins of the Japanese: Common ground for hunter-gatherer and farmer Y chromosomes". Journal of Human Genetics. 51 (1): 47–58. doi:10.1007/s10038-005-0322-0. PMID 16328082.
- Ilumäe (2016). "Human Y Chromosome Haplogroup N: A Non-trivial Time-Resolved Phylogeography that Cuts across Language Families". American Journal of Human Genetics. 99 (1): 163–73. doi:10.1016/j.ajhg.2016.05.025. PMC 5005449. PMID 27392075.
- Kang Hu (2015). "The dichotomy structure of Haplogroup N". arXiv:1504.06463 [q-bio.PE].
- Karafet, Tatiana; Xu, Liping; Du, Ruofu; Wang, William; Feng, Shi; Wells, R.S.; Redd, Alan J.; Zegura, Stephen L.; Hammer, Michael F. (2001). "Paternal Population History of East Asia: Sources, Patterns, and Microevolutionary Processes". The American Journal of Human Genetics. 69 (3): 615–28. doi:10.1086/323299. PMC 1235490. PMID 11481588. In this article, the "Southern Han" sample of Karafet and Hammer's research group is described as originating from Guangdong, and the "Northern Han" sample is described as originating from Shaanxi.
- Karafet, T. M.; Mendez, F. L.; Meilerman, M. B.; Underhill, P. A.; Zegura, S. L.; Hammer, M. F. (2008). "New binary polymorphisms reshape and increase resolution of the human Y chromosomal haplogroup tree". Genome Research. 18 (5): 830–38. doi:10.1101/gr.7172008. PMC 2336805. PMID 18385274.
- Karafet, T. M.; Hallmark, B.; Cox, M. P.; Sudoyo, H.; Downey, S.; Lansing, J. S.; Hammer, M. F. (2010). "Major East-West Division Underlies Y Chromosome Stratification across Indonesia". Molecular Biology and Evolution. 27 (8): 1833–44. doi:10.1093/molbev/msq063. PMID 20207712.
- Katoh, Toru; Munkhbat, Batmunkh; Tounai, Kenichi; Mano, Shuhei; Ando, Harue; Oyungerel, Ganjuur; Chae, Gue-Tae; Han, Huun; Jia, Guan-Jun; Tokunaga, Katsushi; Munkhtuvshin, Namid; Tamiya, Gen; Inoko, Hidetoshi (2005). "Genetic features of Mongolian ethnic groups revealed by Y-chromosomal analysis". Gene. 346: 63–70. doi:10.1016/j.gene.2004.10.023. PMID 15716011.
- Kharkov, V. N.; Stepanov, V. A.; Medvedeva, O. F.; Spiridonova, M. G.; Voevoda, M. I.; Tadinova, V. N.; Puzyrev, V. P. (2007). "Gene pool differences between Northern and Southern Altaians inferred from the data on Y-chromosomal haplogroups". Russian Journal of Genetics. 43 (5): 551–62. doi:10.1134/S1022795407050110.
- Kim, Wook; Yoo, Tag-Keun; Kim, Sung-Joo; Shin, Dong-Jik; Tyler-Smith, Chris; Jin, Han-Jun; Kwak, Kyoung-Don; Kim, Eun-Tak; Bae, Yoon-Sun (2007). Blagosklonny, Mikhail, ed. "Lack of Association between Y-Chromosomal Haplogroups and Prostate Cancer in the Korean Population". PLoS ONE. 2 (1): e172. Bibcode:2007PLoSO...2..172K. doi:10.1371/journal.pone.0000172. PMC 1766463. PMID 17245448.
- Lappalainen, T.; Laitinen, V.; Salmela, E.; Andersen, P.; Huoponen, K.; Savontaus, M.-L.; Lahermo, P. (2008). "Migration Waves to the Baltic Sea Region". Annals of Human Genetics. 72 (3): 337–48. doi:10.1111/j.1469-1809.2007.00429.x. PMID 18294359.
- Malyarchuk, Boris; Derenko, Miroslava; Grzybowski, Tomasz; Lunkina, Arina; Czarny, Jakub; Rychkov, Serge; Morozova, Irina; Denisova, Galina; Miscicka-Sliwka, Danuta (2004). "Differentiation of Mitochondrial DNA and Y Chromosomes in Russian Populations". Human Biology. 76 (6): 877–900. doi:10.1353/hub.2005.0021. PMID 15974299.
- Pakendorf, Brigitte; Morar, Bharti; Tarskaia, Larissa; Kayser, Manfred; Soodyall, Himla; Rodewald, Alexander; Stoneking, Mark (2002). "Y-chromosomal evidence for a strong reduction in male population size of Yakuts". Human Genetics. 110 (2): 198–200. doi:10.1007/s00439-001-0664-4. PMID 11935328.
- Rootsi, Siiri; Zhivotovsky, Lev A; Baldovič, Marian; Kayser, Manfred; Kutuev, Ildus A; Khusainova, Rita; Bermisheva, Marina A; Gubina, Marina; et al. (2006). "A counter-clockwise northern route of the Y-chromosome haplogroup N from Southeast Asia towards Europe". European Journal of Human Genetics. 15 (2): 204–11. doi:10.1038/sj.ejhg.5201748. PMID 17149388.
- Wen, Bo; Xie, Xuanhua; Gao, Song; Li, Hui; Shi, Hong; Song, Xiufeng; Qian, Tingzhi; Xiao, Chunjie; et al. (2004b). "Analyses of Genetic Structure of Tibeto-Burman Populations Reveals Sex-Biased Admixture in Southern Tibeto-Burmans". The American Journal of Human Genetics. 74 (5): 856–65. doi:10.1086/386292. PMC 1181980. PMID 15042512.
- Xue, Y.; Zerjal, T; Bao, W; Zhu, S; Shu, Q; Xu, J; Du, R; Fu, S; et al. (2005). "Male Demography in East Asia: A North-South Contrast in Human Population Expansion Times". Genetics. 172 (4): 2431–39. doi:10.1534/genetics.105.054270. PMC 1456369. PMID 16489223.
Websites
- ISOGG (2012). "Y-DNA Haplogroup Tree 2012".
- McDonald, Doug. "Macdonald Y Haplogroups of the World" (PDF).
Further reading
- ISOGG (2006). "Y-DNA Haplogroup Tree 2006".
- ISOGG (2007). "Y-DNA Haplogroup Tree 2007".
- ISOGG (2008). "Y-DNA Haplogroup Tree 2008".
- ISOGG (2009). "Y-DNA Haplogroup Tree 2009".
- ISOGG (2010). "Y-DNA Haplogroup Tree 2010".
- ISOGG (2011). "Y-DNA Haplogroup Tree 2011".
- ISOGG (2014). "Y-DNA Haplogroup Tree 2014".
- YFull. "YFull Experimental YTree".
Phylogenetics
- ↑ The b2/b3 deletion in the AZFc region of the human Y-chromosome is a characteristic of Haplogroup N-M231 haplotypes. This deletion, however, appears to have occurred independently on four different occasions. Therefore this deletion should not be thought as a unique event polymorphism contributing to the definition of this branch of the Y-chromosome tree (ISOGG 2012).
- ↑ This table shows historic names for N-M46 (AKA N-Tat) from peer reviewed literature.
YCC 2002/2008 (Shorthand) N-M46/N-TAT Jobling and Tyler-Smith 2000 12 Underhill 2000 VIII Hammer 2001 1I Karafet 2001 26 Semino 2000 Eu13 Su 1999 H5 Capelli 2001 F YCC 2002 (Longhand) N3* YCC 2005 (Longhand) N3 YCC 2008 (Longhand) N1c YCC 2010r (Longhand) N1c - ↑ This table shows historic names for N-M178 from peer reviewed literature.
YCC 2002/2008 (Shorthand) N-M178 Jobling and Tyler-Smith 2000 16 Underhill 2000 VIII Hammer 2001 1I Karafet 2001 26 Semino 2000 Eu14 Su 1999 H5 Capelli 2001 F YCC 2002 (Longhand) N3a* YCC 2005 (Longhand) M178 YCC 2008 (Longhand) N1c1 YCC 2010r (Longhand) N1c1 - ↑ This table shows historic names for N-M128 from peer reviewed literature.
YCC 2002/2008 (Shorthand) N-M128 Jobling and Tyler-Smith 2000 12 Underhill 2000 VIII Hammer 2001 1U Karafet 2001 25 Semino 2000 Eu16 Su 1999 H5 Capelli 2001 F YCC 2002 (Longhand) N1 YCC 2005 (Longhand) N1 YCC 2008 (Longhand) N1a YCC 2010r (Longhand) N1a - ↑ This branch is sometimes called N1b in early trees.
External links
| Wikimedia Commons has media related to Haplogroup N of Y-DNA. |
- Spread of Haplogroup N, from The Genographic Project, National Geographic
- N North Eurasian YDNA Project at FamilyTreeDNA
- N Y-DNA Haplogroup Project at FamilyTreeDNA
- N1c1 Y-DNA Haplogroup Project at FamilyTreeDNA
- Y-chromosome haplogroup N dispersals from south Siberia to Europe
- Rurikid Dynasty DNA Project at FamilyTreeDNA
- Russian Nobility DNA Project at FamilyTreeDNA