Hadrosaurus
Hadrosaurus is een plantenetende ornithischische dinosauriër die tot de Euhadrosauria behoorde. De soort Hadrosaurus foulkii leefde tijdens het Laat-Krijt in het gebied van het huidige Noord-Amerika. Het fossiel materiaal van Hadrosaurus is van zulke slechte kwaliteit dat die naam tegenwoordig vaak als een nomen dubium beschouwd wordt: het is niet mogelijk van andere fossielen vast te stellen of ze tot hetzelfde geslacht behoren.
| Hadrosaurus Status: Uitgestorven, als fossiel bekend | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | ||||||||||||||
| Taxonomische indeling | ||||||||||||||
| ||||||||||||||
| Geslacht | ||||||||||||||
| Hadrosaurus Leidy, 1858 | ||||||||||||||
| Typesoort | ||||||||||||||
| Hadrosaurus foulkii | ||||||||||||||
| Afbeeldingen op | ||||||||||||||
| Hadrosaurus op | ||||||||||||||
| ||||||||||||||
Ontdekking en naamgeving
Het eerste skelet van een uitgestorven dinosauriër uit Amerika
Rond 1838 werden grote botten ontdekt door John Estaugh Hopkins, de bezitter van een kleine steengroeve waar mergel gedolven werd voor de verbetering van de structuur van akkerbouwgrond, bij een meertje dat tegenwoordig Hopkins Pond heet, nabij Haddonfield in Camden County, New Jersey. Ten onrechte geven sommige bronnen diens vader William Estaugh Hopkins de eer van de ontdekking. J.E. Hopkins liet die beenderen opstellen in zijn villa Birdwood, eveneens in Haddonfield, een kilometer voor de groeve. In de loop der jaren gaf hij de fossielen alle weg aan bezoekers.

In de zomer van 1858 kreeg een buurman, William Parker Foulke, een natuurvorser uit Philadelphia, tijdens een diner het verhaal over de botten te horen. Foulke, die dacht dat het om beenderen van Mosasaurus moest gaan, spoorde met veel moeite de precieze locatie op en liet hetzelfde jaar de rest van het overgebleven skelet uitgraven aan de rand van de oude steengroeve, bij een beekje dat noordelijk naar de Cooper Creek vloeit. Foulke voerde een voor die tijd nauwgezette documentatie uit waarbij hij de precieze positie van ieder bot in de mergel noteerde. Nadat een bot vrijgelegd was, werd het gestabiliseerd om te voorkomen dat het uit elkaar zou vallen en in stro verpakt naar Foulke's huis vervoerd. Een van de werklui wist een deel van de weggeschonken botten op te sporen. In oktober werden tijdens een tweede opgraving nog wat beenderen gevonden. Uiteindelijk ging het om de achterpoten, een voorpoot, het bekken, achtentwintig wervels, acht tanden en delen van de kaken. De locatie is sinds 1994 een officieel National Historic Landmark, de Hadrosaurus Foulkii Leidy Site. Het beekje heet nu de Hadrosaurus Run. Hadrosaurus is, ondanks zijn dubieuze status, sinds 1994 de officiële staatsdinosauriër van New Jersey.
Foulke kreeg al gauw in de gaten dat het niet om een lid van de Mosasauridae ging maar om een geheel unieke vondst. Hij nam contact op met een gezaghebbende paleontoloog, Joseph Leidy, die in Philadelphia aan de Academy of Natural Sciences werkte als professor anatomie en conservator van het museum van de instelling. Leidy hield een voordracht over het dier op 14 december 1858. In 1858 en 1859 publiceerde hij beschrijvingen. Daarbij benoemde hij in 1858 — in feite via de gedrukte tekst van de voordracht — de typesoort Hadrosaurus foulkii. De geslachtsnaam betekent "forse sauriër" en is afgeleid van het Oudgrieks ἁδρός, hadros, "fors, gezet", een verwijzing naar de omvang van het dier. De soortaanduiding eert Foulke. De beenderen werden aan het museum geschonken wat wel gezien is als een aanwijzing dat de commodificatie van fossielen nog niet ver was voortgeschreden: een generatie later zou voor zo'n zeldzaam exemplaar vermoedelijk een aanzienlijke geldsom zijn gevraagd.

Hadrosaurus was in 1858 vooral opzienbarend omdat het toen de meest compleet bewaard gebleven dinosauriër was — en het eerste dinosauriërskelet uit de Nieuwe Wereld. Binnen tien jaar zouden in het westen van Amerika grote aantallen veel vollediger skeletten gevonden worden maar voor 1858 waren uit dat continent alleen losse botten, tanden en voetafdrukken bekend. Van Hadrosauridae waren er maar twee vondsten gedaan die Leidy zelf in 1856 benoemd had: Trachodon mirabilis die gebaseerd was op wat tanden en Thespesius occidentalis waarvan het materiaal bestond uit twee staartwervels en een kootje. Leidy was daarom gedwongen om vergelijkingen te maken met eerdere Europese vondsten. Hij vond het dier het meest lijken op Iguanodon, iets wat achteraf een correcte waarneming bleek want daaraan is Hadrosaurus inderdaad tamelijk nauw verwant. Overigens zou Hadrosaurus niet lang het meest complete dinosauriërfossiel blijven: in 1859 werd de veel vollediger Compsognathus ontdekt.

Het typespecimen is gevonden in een laag van de Woodburyformatie die dateert uit het Campanien en ongeveer tachtig miljoen jaar oud is. Het wordt vaak aangegeven als het holotype ANSP 10005. In feite echter bestaat het uit een reeks syntypen. ANSP 10005 is slechts het inventarisnummer van het postcraniaal skelet, dus de delen achter de schedel. De gevonden schedelfragmenten hebben eigen nummers. Dat heeft aanleiding gegeven tot het misverstand dat bij het skelet helemaal geen schedeldelen aangetroffen zijn. De syntypen zijn: ANSP 9203: een stuk van de zijwand van het midden van het rechterbovenkaaksbeen; ANSP 9204: een meer naar buiten gelegen stuk van interne vleugel van het linkerbovenkaaksbeen die contact maakte met het ectopterygoïde; ANSP 9202: delen van de tanddragende beenwallen van de kaken die overigens slechts lege tandkassen bevatten; ANSP 9201: een partij losse tanden uit de boven- en onderkaken; en ten slotte ANSP 10005: drie voorste ruggenwervels; negen wervellichamen uit het voorste en middelste deel van de staart, waarbij één middelste staartwervel vrijwel onbeschadigd is; talrijke kleine stukken wervels; een gedeeltelijk rechterravenbeksbeen; een linkeropperarmbeen; een linkerellepijp; een linkerspaakbeen; een linkerdarmbeen; een rechterzitbeen; een stuk rechterschaambeen; en van de linkerachterpoot een dijbeen, een scheenbeen, een vrijwel volledig kuitbeen, een tweede en vierde middenvoetsbeen en het eerste kootje van de derde teen. De Woodburyformatie bestaat uit zeeafzettingen en het kadaver van Hadrosaurus is vermoedelijk door een rivier in zee gespoeld.
Buiten het holotype is een klein aantal losse botten uit het Krijt van de oostkust van Amerika ooit aan Hadrosaurus foulkii toegewezen. Het betreft AMNH 1460: een stuk wervellichaam; AMNH 7626: een stuk rechterdentarium; ANSP 15716: de onderkant van een linkeropperarmbeen; en ANSP 15717: een stuk van een derde middenvoetsbeen. Hoewel het hier om hadrosauride materiaal gaat dat in de laatste twee gevallen inderdaad uit de Woodburyformatie komt, delen de botten geen unieke kenmerken met het holotype en kunnen naar moderne normen dus ook niet aan Hadrosaurus toegewezen worden.
Eerste interpretaties

In 1860 gaf Leidy een gedetailleerdere beschrijving in zijn Cretaceous Reptiles of the United States, welk boek vanwege de Amerikaanse Burgeroorlog pas in 1865 gepubliceerd werd. Leidy beschouwde al in 1858 Hadrosaurus als een tweevoeter, terwijl de dinosauriërs tot dan meestal allen als viervoeters gezien werden. Een uitzondering was Iguanodon waarvan Gideon Mantell vlak voor diens dood begon te vermoeden dat hij op zijn achterpoten liep. Rond 1870 zouden de nieuwe vondsten uit het westen een dergelijke lichaamshouding voor veel dinosauriërs bevestigen. In de jaren zeventig van de twintigste eeuw echter werd duidelijk dat dieren als Hadrosaurus meestal op vier poten rondstapten en zich alleen op de achterpoten verhieven om te rennen. Overigens hield Leidy daar al in 1858 rekening mee en waarschuwde voor het trekken van overhaaste conclusies uit het feit dat de achterpoten zoveel groter waren daar er veel moderne reptielen en amfibieën bestaan met eenzelfde wanverhouding tussen de voorste en achterste ledematen terwijl ze toch "gewoon" op vier poten lopen. Een verschil met Hadrosaurus is wel dat die van een volledig tweevoetige kleinere voorouder afstamde.
%2C_b._1850.jpg)
In 1868 werd er op basis van de vondsten door de beeldhouwer Benjamin Waterhouse Hawkins een opstelling van een skeletmodel gemaakt voor de Philadelphia Academy of Natural Sciences, die tot op heden daar aanwezig is. Dit was de eerste keer dat een Mesozoïsch dinosauriërskelet opgesteld getoond werd, in plaats van als een verzameling losse botten of een steenplaat met fossielen. Het model deed de bezoekersaantallen van de museumafdeling verdriedubbelen. Hadrosaurus werd door Hawkins, conform de ideeën van Leidy, in een opgerichte positie afgebeeld. De constructie van het model hield verband met een veel uitgebreider project. Vanaf 1868 werd namelijk door Hawkins op verzoek van Park Commissioner Andrew Haswell Green een ander volledig skeletmodel en drie modellen van het levende dier — een liggend, een tweede in gevecht met Laelaps, een derde als kadaver waarover twee theropoden vechten — gemaakt voor het Central Park Museum in New York. Behalve Hadrosaurus gaf Hawkins gestalte aan een groot aantal andere prehistorische levensvormen op levende grootte. Dat project werd echter in december 1870 beëindigd door de kliek rond de corrupte politicus William M. Tweed. Toen Hawkins zich in 1871 openlijk tegen Tweed uitsprak, liet die de betonnen beelden op 3 mei door zijn handlanger Henry Hilton kapotslaan. De resten zijn daarop ten dele gebruikt als vulmateriaal en ten dele in The Pond geworpen, de kleinere vijver aan de zuidzijde van Central Park, waarna ze verloren zijn gegaan toen die schoongemaakt werd. Een derde skeletmodel door Hawkins bevindt zich in het Smithsonian Institute maar in een onttakelde toestand, een vierde werd sinds 1879 in het Royal Scottish Museum in Edinburgh opgesteld maar is in 1928 afgedankt en een vijfde bevindt zich in de Nassau Hall van Princeton University. In Philadelphia werd later een moderne reconstructie van het skelet tentoongesteld; het oude model bevindt zich meestal in opslag. Daarnaast toont men er de oorspronkelijke botten op een silhouet van het dier. Pas in juli 1883 werd een tweede Mesozoïsch dinosauriërskelet opgesteld, dat van Iguanodon te Brussel. Hierbij ging het om het fossiel zelf.
Latere interpretaties
Hadrosaurus was tot het eind van de negentiende eeuw de enige hadrosauride waarvan wat ruimer materiaal bekend was. Verschillende geleerden besteedden daarom aandacht aan de soort. In 1868 werd het materiaal opnieuw beschreven door Edward Drinker Cope. In 1869 gaf Cope een volgende, nog gedetailleerdere, beschrijving. Daarbij gaf hij ook een reconstructie van het bekken, waarin een door Leidy fout geïdentificeerd bot correct als een deel van wat het schaambeen was, werd onderkend. Cope maakte echter de fout het schaambeen en het zitbeen te verwisselen. Daarnaast meldde hij een stuk van de onderkant van het schouderblad ontdekt te hebben. Dit kon door latere onderzoekers niet gelokaliseerd worden.
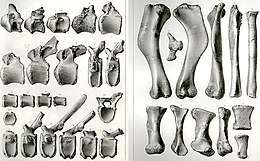
Cope stelde in 1869 Trachodon en Thespesius aan Hadrosaurus gelijk, vergetend dat die genera dan prioriteit zouden hebben. Hoewel Leidy zelf in 1868 ook aan een identiteit gedacht had, verwierp hij die in 1870. In 1874 dacht Cope dat ook Claosaurus aan Hadrosaurus identiek was. Datzelfde jaar en in 1875 kwam Hawkins op zijn restauraties terug en onderkende dat elementen die Leidy aangezien had voor pseudo-clavicles, dus botten die in positie overeenkwamen met de sleutelbeenderen maar waarvan hij dacht dat die dit toch niet konden zijn omdat hij ten onrechte aannam dat die bij dinosauriërs ontbraken, in feite delen van het bekken waren, van het zitbeen. Hawkins was in 1875 ook degene die Cope's verwisseling van schaambeen en zitbeen ontdekte, die hij tot zijn spijt in zijn reconstructies gevolgd had in plaats van Leidys oorspronkelijke correcte identificatie. In 1888 stelde Richard Lydekker Hadrosaurus aan Trachodon gelijk.

In de twintigste eeuw was de belangstelling voor Hadrosaurus een stuk minder. Rond 1900 was een groot aantal hadrosauriden ontdekt met vrijwel complete fossielen en deze trokken meer de aandacht dan een onvolledige vorm die alleen nog een historisch belang leek te hebben. In 1942 werd Hadrosaurus nog eens beschreven door Richard Swann Lull en Nelda Wright. Deze vergeleken de soort vooral met Anatosaurus en Edmontosaurus. Ze zagen veel overeenkomsten maar beschouwden deze laatste geslachten toch als verschillend van Hadrosaurus. In 1977 stelden Donald Baird en Jack Horner dat Gryposaurus — die indertijd gemeenlijk bij Kritosaurus ondergebracht werd — een jonger synoniem was van het geslacht Hadrosaurus, ondanks een aanzienlijk verschil in leeftijd en herkomst terwijl ze de stelling niet ondersteunden door een anatomische vergelijking. In de jaren daarna werd veel aandacht besteed aan deze hypothese, waarbij de conclusie of was dat de taxa niet nauw verwant waren of dat ze wel als nauw verwant konden gelden maar dan toch verschillend. Horner zelf nam in 1990 deze laatste positie in. Het probleem wordt gecompliceerd doordat er tegenwoordig drie soorten binnen Gryposaurus worden onderscheiden. Hadrosaurus foulkii kan niet gelijk zijn aan G. latidens daar die laatste vorm, zoals zijn soortaanduiding al aangeeft, veel bredere tanden heeft en daarbij een hoger voorblad van het darmbeen. In 2006 concludeerde een studie dat alle soorten van Gryposaurus, hoewel in de proporties van de ledematen sterk met Hadrosaurus overeenkomend toch daarvan verschillen door een langere en hoekiger deltopectorale kam op het opperarmbeen; daarbij kromt hun zitbeen typisch omlaag in plaats van omhoog.
Hoewel Hadrosaurus in 1858 uitzonderlijk volledig bewaard was, werd het uiteindelijk een relatief slecht bekende vorm. Later werden er namelijk vele andere soorten in de Hadrosauridae ontdekt en beschreven. Hierdoor werd de slechte conservering van het fossiel en het ontbreken van het grootste deel van de schedel een probleem voor de geldigheid van de soort. De Hadrosauridae lijken erg op elkaar in de postcrania, de delen van het skelet achter de schedel; als deze laatste niet bewaard is gebleven, kan slechts aan kleine details worden gezien tot welke soort een fossiel behoort — en deze details ontbreken bij het holotype van Hadrosaurus. Eerdere onderzoekers maakten daar niet zo'n probleem van, ook omdat ze meer belang hechtten aan de unieke ouderdom en herkomst. Tegen het eind van de twintigste eeuw echter werd steeds meer de klemtoon gelegd op de verzameling unieke kenmerken die iedere soort moet bezitten, ook omdat daarmee in de methode van de cladistiek de positie in de stamboom werd bepaald. In 2006 concludeerden Albert Prieto-Márquez en David Weishampel dat Hadrosaurus helemaal geen bepaalbare unieke kenmerken had en dus een nomen dubium was, een twijfelachtige naam waaraan geen andere fossielen gerechtvaardigd kunnen worden toegewezen. Verschillende andere soorten die in het geslacht waren benoemd: H. tripos, H. minor, H. cavatus, H. breviceps en H. paucidens werden ook als nomina dubia afgewezen. In 2011 veranderde Prieto-Márquez van opvatting en dacht dat Hadrosaurus toch geldig was. Sommige andere onderzoekers echter blijven erbij dat Hadrosaurus een nomen dubium is.
Verdere soorten
Net als bij andere vroeg benoemde dinosauriërs, zoals Iguanodon en Megalosaurus, werden aan het geslacht Hadrosaurus verschillende slecht bekende vormen toegeschreven. Het geslacht fungeerde zo als "prullenmand" voor allerlei onduidelijk hadrosauride — of vermeend hadrosauride — materiaal.
Leidy zelf hernoemde in 1868 zijn Trachodon mirabilis in een Hadrosaurus mirabilis.
In 1869 benoemde Cope een Hadrosaurus tripos, de "drievoet", gebaseerd op de specimina USNM 7190, 7093, 7094 en 7095, vier staartwervels gevonden in de mergelgroeve van W.J. Thompson, Sampson County, North Carolina. De groeve exploiteerde een laag van de Duplimergel die dateert uit het Plioceen wat al aantoont dat het niet om een Mesozoïsche dinosauriër kan gaan. In 1979 stelden Horner & Baird vast dat het een baleinwalvis betrof uit de Balaenopteridae en daarbij is het een nomen dubium. Ook in 1869 benoemde Cope een Ornithotarsus immanis gebaseerd op een linkeronderbeen van een zeer grote hadrosauride, specimen YPM 3221. Sommige latere auteurs zagen dit als een jonger synoniem van Hadrosaurus foulkii. Lull & Wright toonden echter aan dat het gaat om een nomen dubium. Hawkins baseerde een deel van zijn reconstructie van de enkel van Hadrosaurus op Ornithotarsus.
In 1870/1871 benoemde professor Othniel Charles Marsh een Hadrosaurus minor, "de kleinere". Het holotype was specimen YPM 1600, een partij wervels van de rug of de nek die waren gevonden bij Barnsboro, Gloucester County, New Jersey, in lagen van de New Egypt Formation die dateert uit het Maastrichtien. Talrijke andere fragmentarische vondsten van en ledematen uit dezelfde formatie, of uit lagen aan de oostkust waarvan Marsh vermoedde dat ze even oud waren, werden aan de soort toegewezen. Het gaat vermoedelijk om resten van hadrosauride jongen. In 1948 wees Edwin Harris Colbert een fragmentarisch skelet van een jong dier, specimen ANSP 15202, dat bij Sewell gevonden was, aan Hadrosaurus minor toe, wegens de grootte en herkomst. De meeste latere onderzoekers achtten de soort een nomen dubium; de wervels van het holotype hebben geen diagnostische kenmerken. Bij ANSP 15202 gaat het mogelijkerwijs om een jong van Edmontosaurus annectens. Het exemplaar lijkt te tonen dat de centra en wervelbogen van de wervels van de staart eerder vergroeiden dan die van de rug, een ontwikkelingsvolgorde die ook van krokodillen bekend is.
In 1871 benoemde Cope Hadrosaurus cavatus, "de uitgeholde", gebaseerd op holotype AMNH 1390, vier staartwervels, wellicht gevonden bij Swedesboro in de New Egypt Formation. Naar moderne inzichten is het een nomen dubium.
In 1872 benoemde Marsh een Hadrosaurus agilis, "de wendbare". In 1890 maakte hij daar het aparte geslacht Claosaurus van. In 1889 benoemde Marsh een ?Hadrosaurus breviceps, "kortekaak", gebaseerd op holotype YPM 1779 gevonden bij Bearpaw Mountain, Yellowstone, Wyoming, uit de Judith River Formation daterend uit het Campanien, een stuk achterste dentarium van de onderkaak met zevenentwintig tandposities. Het vraagteken geeft aan dat Marsh zelf niet overtuigd was van de juistheid van de plaatsing en er slechts voor koos vanuit de "prullemandfunctie". Het stuk onderscheidt zich door een opvallende breedte en een plat in plaats van hol slijtvlak van de tandbatterij. Prieto-Márquez achtte het in 2006 niet onderscheidend genoeg en oordeelde dat het een nomen dubium is. In 1942 hernoemde Lull het tot een Kritosaurus breviceps. Later in 1889 benoemde Marsh een Hadrosaurus paucidens, "kleinetand", gebaseerd op holotype USNM 5457, twee stukken uit de linkerhelft van een schedel, het squamosum en het bovenkaaksbeen, gevonden bij de Dog Creek, Fergus County, Montana uit de Judith River Formation. In 1890 meende Marsh dat hij zich vergist had en het om een lid van de Ceratopia ging — Oliver Perry Hay hernoemde het in 1901 tot een Ceratops paucidens — maar daarin was hij abuis. In 1964 toonde John Harold Ostrom aan dat het een lambeosaurine betrof die hij hernoemde tot Lambeosaurus paucidens. Prieto-Márquez vond in 2006 dat het een nomen dubium was, een niet verder te determineren lid van de Lambeosaurinae.
In 1892 hernoemde Edwin Tulley Newton ?Trachodon cantabrigiensis Lydekker 1888 in een Hadrosaurus cantabrigiensis. De soort is gebaseerd op een tand, specimen BMNH R496, gevonden bij Cambridge.
In 1900 hernoemde baron Franz Nopcsa Thespesius occidentalis in een Hadrosaurus occidentalis — waarvan hij ook nog meende dat Agathaumas milo een jonger synoniem was — en Trachodon longiceps Marsh 1890 in een Hadrosaurus longiceps.
In 1901 hernoemde Hay Agathaumas milo Cope 1874 in een Hadrosaurus milo.
In 1977 hernoemden Donald Baird en Jack Horner Gryposaurus notabilis Lambe 1914 in een Hadrosaurus notabilis en Kritosaurus navajovius Brown 1910 in een Hadrosaurus navajovius.
Soortenlijst
De ingewikkelde naamgevingsgeschiedenis kan worden samengevat in een soortenlijst.
- Hadrosaurus foulkii Leidy 1858: typesoort van Hadrosaurus, wellicht nomen dubium; wellicht ouder synoniem van Ornithotarsus Cope 1869; = Trachodon foulkii (Leidy 1858) Lydekker 1888
- Hadrosaurus mirabilis (Leidy 1856) Leidy 1868: nomen dubium; = Trachodon mirabilis Leidy 1856
- Hadrosaurus tripos Cope 1869: nomen dubium, non Hadrosaurus, een baleinwalvis
- Hadrosaurus minor Marsh 1870: nomen dubium; = Edmontosaurus minor Baird & Horner 1977
- Hadrosaurus cavatus Cope 1871: nomen dubium, = Trachodon cavatus (Cope 1871) Hay 1902
- Hadrosaurus agilis Marsh 1872: = Claosaurus agilis (Marsh 1872) Marsh 1890; = Trachodon agilis (Marsh 1872) Kuhn 1936
- ?Hadrosaurus breviceps Marsh 1889: nomen dubium; = Trachodon breviceps (Marsh 1889) Hay 1902; = Kritosaurus breviceps (Marsh 1889) Lull & Wright 1942
- Hadrosaurus paucidens Marsh 1889: nomen dubium; = Ceratops paucidens (Marsh 1889) Hay 1901
- Hadrosaurus cantabrigiensis (Lydekker 1888): nomen dubium; = ?Trachodon cantabrigiensis Lydekker 1888; = Telmatosaurus cantabrigiensis (Lydekker 1888) Olshevsky 1978
- Hadrosaurus occidentalis (Leidy 1856) Nopcsa 1900: nomen dubium; = Thespesius occidentalis Leidy 1856; = Trachodon occidentalis (Leidy 1856) Kuhn 1936
- Hadrosaurus longiceps (Marsh 1890) Nopcsa 1900: = Trachodon longiceps Marsh 1890; = Anatotitan longiceps (Marsh 1890) Olshevsky 1991
- Hadrosaurus milo (Cope 1874) Hay 1901: nomen dubium; = Agathaumas milo Cope 1874
- Hadrosaurus notabilis (Lambe 1914) Baird & Horner 1977: = Gryposaurus notabilis Lambe 1914; = Kritosaurus notabilis (Lambe 1914) Lull & Wright 1942
- Hadrosaurus navajovius (Brown 1910) Baird & Horner 1977: = Kritosaurus navajovius Brown 1910
Beschrijving

Grootte
Het individu van het holotype van Hadrosaurus was zo'n achtenhalve meter lang. Dat wijst op een gewicht van ruim drie ton. Leidy schatte in 1858 de lengte op acht meter. Cope stelde die in 1868 op vierentwintig tot dertig voet, dus 8,4 tot negen meter. Gregory S. Paul kwam in 2010 op een kleinere lengte uit van zeven meter. Sommige bronnen meldden veel grotere lichaamslengten, tot wel vijftien meter, maar die schattingen zijn gebaseerd op Ornithotarsus.
Onderscheidende kenmerken
Volgens wetenschappers die Hadrosaurus als een nomen dubium beschouwen had het dier dus geen onderscheidende kenmerken. Albert Prieto-Márquez was in 2006 die mening toegedaan. In 2011 veranderde hij van mening en stelde dat Hadrosaurus althans een unieke combinatie van op zich niet unieke kenmerken toonde en dus een geldig taxon was. Die kenmerken bevonden zich in het bekken en de ledematen. Opvallend vond hij vooral de combinatie van een korte deltopectorale kam op het opperarmbeen, de hoge richel aan de voorkant waaraan belangrijke spieren vastzitten die dat bot bewogen, met een brede gewelfde buitenste onderhoek van diezelfde kam. Deze kenmerken zijn basaal, dat wil zeggen dat ze ook bij eerdere Iguanodontia voorkomen.
Daarnaast waren er afgeleide kenmerken, dus eigenschappen die ook voorkomen bij verwanten die hoger in de stamboom staan, in dit geval Edmontosaurus en Shantungosaurus. De antitrochanter, een buitenste uitstulping van het bovenste heupgewricht, van het darmbeen is kort in zijaanzicht. De buitenste onderrand van de antitrochanter, op halve hoogte van het darmbeenblad, is symmetrisch U-vormig. De achterste onderrand van de antitrochanter, waar die het achterblad van het darmbeen raakt, is slecht ontwikkeld. Het bovendeel van het zitbeen, dat contact maakt met het darmbeen, kromt niet naar achteren. De voorste bovenhoek van dat bovendeel is verheven ten opzichte van de schacht van het zitbeen.
De mengeling van basale en afgeleide kenmerken was volgens Prieto-Márquez uniek. In 2014 wezen andere onderzoekers er echter op dat de vermeende kortheid van de deltopectorale kam, minder dan 48% van de schachtlengte, juist veroorzaakt wordt door de gebogen onderhoek ervan en gezien moet worden als een geval van individuele variatie. Ze concludeerden dat Hadrosaurus toch een nomen dubium was.
Fylogenie
Leidy plaatste het dier in 1860 in de Sauria. In 1869 benoemde Cope een eigen familie Hadrosauridae waarvan Hadrosaurus het typegenus is. Lull en Wright waren in 1942 de eersten die onderkenden dat Hadrosaurus althans in bouw het meest verwant is aan vormen die tegenwoordig de Hadrosaurinae worden genoemd, dieren met vrij platte koppen. In 1961 werd dit echter betwijfeld door John Harold Ostrom. Hij vreesde dat het fragmentarische karakter van de fossielen uitsloot dat ooit met enige zekerheid zou kunnen worden bepaald of Hadrosaurus een lid is van de Hadrosaurinae of een andere subgroep. Hij begreep ook hoe dit zou kunnen leiden tot grote taxonomische instabiliteit daar de Hadrosaurinae dan immers hernoemd zouden moeten worden. Ter wille van de stabiliteit bleef hij Hadrosaurus maar onder de hadrosaurinen scharen.
Horner meende in 2004 dat Hadrosaurus een bepaalde ontwikkelingsgraad vertegenwoordigde, basaal in de Euhadrosauria. Deze graad zou zich onder andere kenmerken door een vermindering van de tandgrootte en het voorkomen van een enkele goed ontwikkelde rand op de dentaire tanden.
Levenswijze
Leidy speculeerde al in 1858 over de levenswijze van het dier. Uit het feit dat het zich op twee poten kon oprichten, leidde hij af dat het als een kangoeroe op zijn staart rustte om hoger gebladerte te eten. Hij dacht echter ook dat Hadrosaurus vaak het, zoete, water opzocht. Leidy begreep al in 1865 uit de vorm van de tanden dat die nauw aaneengesloten moesten zijn geweest — hoewel hij de typische tandbatterijen van de Hadrosauridae nog niet kon kennen.
Cope meende dat het dier aan land op twee poten liep, met gebogen knieën en de staart over de grond slepend. Hij zag het typische leefgebied als zoutmoerassen maar dacht net als Leidy dat Hadrosaurus zich oprichtte om hogere plantenlagen te bereiken, waaraan hij het detail toevoegde dat hoge takken met de voorpoten naar de bek getrokken werden. Ervan uitgaande dat vlezige wangen ontbraken, nam hij aan dat "de ontbloting van verschillende rijen schijnende tanden een ietwat grijnzende fysiognomie verschaft zouden hebben". In 1883 was Cope tot de conclusie gekomen dat het gebit niet toestond hard voedsel te eten. Hij dacht dat zachte waterplanten verorberd werden of anders vissen zonder schubben.
Naar moderne inzichten liep Hadrosaurus meestal op vier poten. Zich oprichten op de staart zal lastig geweest zijn daar die met verbeende pezen verstijfd was en recht naar achteren stak. Het dier was nauwelijks aangepast aan een zwemmende levenswijze en in feite een typische landbewoner. Zachte waterplanten zullen zelden op het dieet hebben gestaan. Het voedsel moet bestaan hebben uit vrij harde lage begroeiing. Hadrosaurus had weinig moeite die te verwerken. Met de scherpe hoornsnavel konden flinke stukken worden afgebeten. Die werden verder verknipt en vermalen door de tandbatterijen in de boven- en onderkaken. De ruitvormige tanden vormden een aaneengesloten rij waarvan het afgesleten kauwvlak een constant scherpe snijrand verschafte. Aan de binnenste onderkant groeiden steeds nieuwe rijen vervangingstanden aan waarbij de punten van de nieuwe rij perfect tussen de kronen van de bovenliggende rij pasten. Tegelijkertijd kunnen zo, afgaande op de toestand bij verwante soorten, tot wel een zes rijen aanwezig geweest zijn. Mogelijkerwijs werd het voedsel verder vermalen door maagstenen ofwel gastrolieten. In de grote buikholte werd het verder door bacteriën verteerd.
Literatuur
|