Neanderthals in Southwest Asia
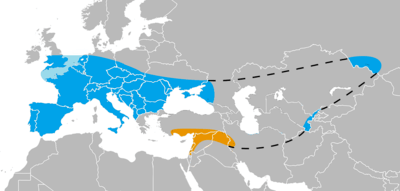
Southwest Asian Neanderthals are Neanderthals that lived in Turkey, Lebanon, Israel, Syria, Iraq, and Iran, the southernmost expanse of their known range. Although their arrival in Asia is not well-dated, early Neanderthals occupied the region apparently until about 100,000 years ago. At this time, Homo sapiens seems to have replaced them in one of the first anatomically modern expansions out of Africa. In their turn, starting around 80,000 years ago, Neanderthals seem to have replaced Homo sapiens in Southwest Asia. They inhabited the region until about 55,000 years ago.[1]
In Southwest Asia, Neanderthals left well-preserved skeletal remains in Israel, Syria, and Iraq. Remains in Turkey, Lebanon, and Iran are fragmentary. No Neanderthal skeletal remain has ever been found to the south of Jerusalem, and although there are Middle Palaeolithic Levallois points in Jordan and in the Arabian peninsula, it is unclear whether these were made by Neanderthals or anatomically modern humans.[1] Neanderthals further to the east, such as those found in Uzbekistan[2][3] and Asian Russia[4][5] are known as Central and North Asian Neanderthals.
As of 2013, although many more Neanderthal remains have been discovered in Southwest Asia than in North Asia, where genetic studies have succeeded,[6] no attempt at extracting DNA from Southwest Asian Neanderthals has ever been successful.[7]
Early hominins (to 100 ka)
As the Levant is the landbridge to Eurasia, Dmanisi remains in Georgia from 1.81 Ma suggest that hominins passed through the Levant some time before this (unless they crossed the Bab el-Mandeb strait into Arabia). The oldest known hominin specimen from the Levant, and from all the Middle East, is the Zuttiyeh skull, found by Francis Turville-Petre near the Sea of Galilee in 1927. It has not been dated, but lithic industries suggest it dates from 250-350 ka (Bar-Yosef 1992).[8] Though it has some modern characteristics, it has been classified as a specimen of Homo heidelbergensis.
Other known hominin fossils in Southwest Asia older than 100 ka include Shanidar 2 and 4, as well as Tabun C1. All three have at one point been classified as Neanderthals, although the latter two are now usually called pre-Neanderthals. In part because none are well-dated these attributions have all been challenged.
Early AMHs in the Levant (100 to 80 ka)
It is believed that anatomically modern humans first entered the Levant by 130 – 100 ka (Grün 2005)[9] from Africa during the Last Interglacial, a hot, parched, period in Africa (Scholz et al. 2007).
Environment
Goat δ13C values from c. 92 ka collected at Qafzeh Cave show that goats were eating, depending on the season, between 20 and 50% of C4 plants. These plants are better adapted to dry and hot climates than the much more common C3 plants. Although it was already established that North Africa was arid in the early Last Glacial, Scholz et al. (2007)'s conclusions would extend this to the Levant, making the region in this period more arid than previously thought. Frumkin et al. (2011)[10] arrived at a similar conclusion of a hot and arid Levant by examining changes through time of lake levels and in the formation of cave formations.
Return of Neanderthals to the Levant (80 to 55 ka)
Potential causes for the initial extinction of modern humans from the Levant and subsequent replacement by Neanderthals around 80 ka include a short-term extreme environmental change in the Levant and environmental changes in the regions neighbouring the Levant. The only long-term environmental change that has been found in the Levant at this period is an increase in rainfall (Hallin et al. 2012),[11] and some moderate cooling of the region (Bar-Yosef 1992,[8] Harrison and Bates 1991),[12] which would not predict a hominin extinction (Hallin et al. 2012).[11] Hence, a short-lived intense environmental change (such as a volcanic winter of 6 to 1000 years caused by the Toba supereruption, Rampino and Self 1992)[13] might have cause modern human extinction, especially if the modern human Levantine population was low (Shea 2008).[14] In this case, Neanderthals could have moved in during or after the modern human evacuation of the region.
Alternatively, climate change could have connected the Neanderthal biome to the north, allowing Neanderthals to replace modern humans if Neanderthals were better pre-adapted to the new Levantine environment. It is also possible for climate change to have allowed the North African desert to expand and cut off the modern human Levantine biome from the larger African one. This could have driven to extinction the modern human population with too few individuals to survive when disconnected from Africa, allowing Neanderthals to walk into the region. This could have happened even if modern humans were better adapted to the region than Neanderthals. In other words, in this case, the only advantage Neanderthals would have had in the Levant is that they could actually get there.
Environment
Contrarily to modern humans who arrived in the Levant in period particularly warm in both Eurasia and Africa, Neanderthals are thought to have entered the Levant in a period of increasing Eurasian coldness and aridity known as the Last Glacial Period (Tchernov 1989).[15] In the Levant, however, the conditions would have been wetter than those of today and of the preceding interglacial. This is what Hallin et al. (2012)[11] concluded in their study of the δ13C and δ18O[Note 1] values of goat and gazelle enamel from Amud cave in Neanderthals layers dated to 70-53 ka (Glacial Period), after comparing them to values from Qafzeh cave in a modern human layer dated to 92 ka. They found that at Amud, neither the goats nor the gazelle consumed arid-adapted C4 plants, indicating that the environment was not as dry as today. They found the magnitude and pattern of δ18O values in gazelle and goat teeth enamel to indicate that, unlike today, rain fell throughout the year at Amud in the Neanderthal period. Indeed, the variation in δ18O values in the oldest layers of the teeth, formed when the animal is only a few months old, suggested that goats and gazelles were born throughout the year, which today happens only in the Levant in wetter-than-normal conditions.
Belmaker et al. (2011)[16] compared the prevalence in caves of microfaunal bones brought back by Barn and Eagle owls, which are known to accumulate large quantities of rodent skeletons. Since owls are opportunistic hunters and will eat any small mammal, the bone assemblage they leave behind are thought to be accurate indicators of the environment, given that rodents, like beetles, are susceptible to small changes in temperature and humidity. The researchers did find changes in microfaunal bone assemblages, but all were expected by taphonomic bias. They therefore concluded that there were no significant changes in the environment of the Levant between 70 and 55 ka.
Return of AMHs to the Levant (after 55 ka)
Unless modern humans started expanding into Eurasia through Arabia (where stone tools but very few human bones have been found), the Levant is most likely the first ground Neanderthals have lost in the face of expanding modern humans. The youngest Neanderthal specimen in the Levant is about 45,000 years old. From there on Neanderthals seem to have retreated all the way to the Atlantic over a period of about 2500 to 5000 years. Neanderthals would then have gone extinct, as there seems to be no evidence of Neanderthals anywhere in their range after 40,000 BP.
It is unclear why Neanderthals abandoned the Levant or lost it to modern humans. Shea (2008)[14] has argued that Neanderthals were climatically forced out of the region (as modern humans probably were about 75,000 years ago). This is contested. Belmaker et al. (2011)[16] have found that the proportions of microfaunal bones in the Levant between 70 and 55 ka seem stable, despite the fact that small fauna are sensitive to slight changes in temperature or humidity. There would be hence little reason to believe in a slow deterioration of the Neanderthal habitat driving them out by 45 ka. It is possible that climate change drove Neanderthals to extinction quickly some time after 55,000 BP, but Hallin et al. (2012)[11] found strong consistency through time in the δ13C and δ18O values in the teeth of both goats and gazelles, suggesting that the climate was stable in the Levant at this crucial period when Middle Palaeolithic technologies of Neanderthals and early modern humans were replaced by Upper Palaeolithic technology of later modern humans.
Although the environment at this period in the Levant seems to have been stable, Shea (2008)[14] suggested that a rapidly changing environment at the border between the Levant and the Sinai could have stimulated the development of the first Upper Palaeolithic technologies. In this scenario, modern humans would only have been able to displace Neanderthals once they had developed this technology. The advanced tools found in Europe in the millennia thereafter would have developed there and then.
Resource gathering strategy
It is believed that Neanderthals and modern humans in the Levant had different resource gathering behaviours. Lieberman and Shea (1994)[17] analysed their lithic hunting technologies and found their differences to suggest Neanderthals hunted more frequently than modern humans. They argue that Neanderthals would have used their intensive hunting strategy to cope with a biodepleted environment, much like the Inuit of the 20th century used to depend largely on hunting. Henry et al. (2017)[18] found that in the Levant the site exploitation territory of Neanderthals was narrow, as they limited themselves to rugged woodlands, while modern humans of the Upper Palaeolithic would have been generalists with large exploitation territories in both the rugged woodlands and levelled steppe landscapes.
Although it does seem as if the environment favoured the initial success and eventual demise of Neanderthals, it is still unclear whether differences in their resource gathering strategies reflect differences in the environment, in anatomy, or in intelligence.
List of Southwest Asian Neanderthals
The remains of more than 70 Southwest Asian Neanderthals have been found. As of 2017, this list may be considered essentially complete. Sites are sorted west to east, first by country (westernmost site) then within countries. Important remains are in bold.
| Present-day country (country of discovery) | Site | Principal Neanderthal finds | MNI | Geological age (ka) | Descriptions | Notes |
|---|---|---|---|---|---|---|
| Turkey | Karain | Four teeth | 1 | — | Senyürek (1949)[19][20] | |
| Lebanon | Ksâr 'Akil | K2: Teeth and partial maxilla | 1 | — | Ewing (1963)[22] | Ewing lost this specimen while transferring Ksar Akil material from Boston College to Fordham University.[23] |
| Lebanon | El Masloukh | Upper second molar[24] | (1) | — | ? | Neanderthal attribution is stratigraphic, not morphological.[25] |
| Israel | Kebara | KMH1: 7-9 mo. old partial skel.
KMH2: Post-cranial adult ♂ Various fragments KMH3: Milk tooth (m1-r)[26] |
21 + (10) | 64-59[29][30] | KMH1: Smith et al. (1977)[31]
KMH2: Arensburg et al. (1985)[32] |
Neanderthal attribution uncertain in KMH18-23, 25, 29, and 31[26][28] |
| West Bank (Mandatory Palestine) | Shuqba | S-D1: Tooth and cranial frags.[24] | 1 | — | Keith (1931)[33] | |
| Israel (Mandatory Palestine) | Tabun | T C1: Nearly complete adult ♀ T C2: Toothed mandible missing I1 (♂) Various fragments T E1: Right femur shaft (♂?) |
15 | ≈170-90 | McCown (1936) McCown and Keith (1939) |
T C1: Neanderthal attribution is not universally accepted.[37]
As of 1975, the whereabouts of T BC2, B3, and BC6 are unknown.[24]:146 |
| Israel | Ein Qashish | (EQH-2: Third molar)
EQH-3: Adult lower limbs |
1 + (1)[1] | 70-60[1] | Been et al. (2017)[1] | Discovered in 2013, these were the first diagnostically Neanderthal remains in Southwest Asia not found in a cave.[1]
EQH-2: 70% posterior probability that Neanderthal attribution is correct.[1] |
| Israel | Shovakh | (Tooth, M(3)-l[38][Note 2]) | (1) | — | S. Binford (1966)[39] |
"[A]lthough within archaic and modern human ranges of variation, this complex occlusal morphology may suggest that it is more likely to have derived from a Neandertal than an early modern human". (Trinkaus 1987)[38] |
| Israel | Amud | A1: Adult full skeleton ♂
A2: Maxillary fragment A7: 10-mo.-old partial skel. |
3[Note 3][40] | 61-53[40] | A1: Suzuki et al. (1970)[41]
A7: Rak et al. (1994)[42] |
|
| Syria | Dederiyeh | D1: 19-30-month-old full skel.
D2: 21-30-month-old full skel. |
17 | — | D1: Akazawa et al. (1993)[43]
D2: Akazawa et al. (1999) [44] |
|
| Iraq | Shanidar | S1: Adult partial skel. ♂ S2: Adult crushed skel. ♂ |
10 | S2, S4: > 100
Others: 60 |
S1: Stewart (1959)[45] S2: Stewart (1961)[46] |
Shanidar 2 and 4 are sometimes not treated as Neanderthals.
All but Shanidar 3 and 10 (and fragments of 5 excavated in 2015-2016)[50] may have been destroyed in the 2003 invasion of Iraq.[54] |
| Iran | Bisitun | Adult radius shaft | 1 | — | Trinkaus (2006)[55] | |
| Total | 70 + (13) |
- ↑ δ13C and δ18O are so-called isotopic signatures. They respectively measure the prevalence of the heavier carbon-13 (13C) and oxygen-18 (18O) isotopes over the more common carbon-12 and oxygen-16. C4-pathway plants have relatively more 13C (higher δ13C) than C3-pathway plants. Because C4 plants are better adapted to hot and dry climates than C3 plants, high δ13C values are associated with hot and dry environments. To the contrary, a greater prevalence of 18O (higher δ18O) in (land) animal bones reflects a colder environment: Because 16O is lighter than 18O, water molecules of the form H216O evaporate slightly more readily than those of the H218O form. This difference in evaporation is greater the colder the environment gets, so that rain and snow in colder regions have a lower δ18O value. Animals incorporate the δ13C and δ18O information of their environment and it is possible to extract it from their bones and teeth.
- ↑ M?-r according to S. Binford (1966)[39].
- ↑ Valladas, Mercier, and Froget (1999) write that "[o]f the remains of 18 individuals recovered at the Amud Cave, 15 were derived from unambiguous Middle Palaeolithic contexts, all of them located in the northern area of the excavation (Hovers et al., 1995). The stratigraphic distribution of these remains encompasses the layers B1 and B2, with only a single specimen derived from layer B4 (Sakura, 1970). Three individuals bear diagnostic characteristics which define them as Neanderthals. Amud I, the skeleton of an adult male was found at the top of layer B1, while the partial skeleton of the baby Amud 7 (Rak, Kimbet & Hovers, 1994) was recovered from the top of layer B2, just under the contact with the base of layer B1. Amud II, represented by a fragment of the right maxilla, was excavated from layer B2."
See also
- Dawn of Humanity (2015 PBS documentary)
- Gibraltar Neanderthals
- Human timeline
- Life timeline
- Nature timeline
- Origins of Us (2011 BBC documentary)
- Prehistoric Autopsy (2012 BBC documentary)
- The Incredible Human Journey (2009 BBC documentary)
Notes
References
- 1 2 3 4 5 6 7 E. Been; E. Hovers; R. Ekshtain; A. Malinski-Buller; N. Agha; A. Barash; D.E. Bar-Yosef Mayer; S. Benazzi; J.-J. Hublin; L. Levin; et al. (2017). "The first Neanderthal remains from an open-air Middle Palaeolithic site in the Levant". Scientific Reports. 7 (2958). doi:10.1038/s41598-017-03025-z.

- ↑ A. Okladnikov (1949). "Исследование мустьерской стоянки и погребеня неандертальца в гроте Тешик-Таш, Южный Узбекистан (Средняя Азия)" [Investigation of the Mousterian site and burial of a Neanderthal in the cave of Teshik-Tash, Southern Uzbekistan (Central Asia)]. Тешик-Таш. Палеолитический человек (in Russian). Moscow. pp. 7–85.
- ↑ Michelle Glantz; Bence Viola; Patrick Wrinn; Tatiana Chikisheva; Anatoly Derevianko; Andrei Krivoshapkin; Uktur Islamov; Rustam Suleimanov; Terrence Ritzman (2008). "New hominin remains from Uzbekistan". Journal of Human Evolution. 55 (2): 223–237.
- ↑ M.B. Mednikova (2011). "A proximal pedal phalanx of a Paleolithic hominin from Denisova Cave, Altai". Archaeology, Ethnology and Anthropology of Eurasia. 39 (1): 129–138.
- ↑ B. Viola; S.V. Markin; A.P. Buzhilova; M.B. Mednikova; A. Le Cabec; M.V. Shunkov; et al. "New Neanderthal remains from Chagyrskaya Cave (Altai Mountains, Russian Federation)". American Journal of Physical Anthropology. 147 (Suppl. 54): 293–294. Retrieved 11 June 2017.
- ↑ K. Prüfer; F. Racimo; N. Patterson; F. Jay; S. Sankararaman; S. Sawyer; A. Heinze; G. Renaud; P.H. Sudmant; C. de Filippo; et al. (2014). "The complete genome sequence of a Neanderthal from the Altai Mountains". Nature. 505: 43–49. doi:10.1038/nature12886. PMC 4031459. PMID 24352235. (subscription required)
- ↑ J. Hawks (2013). "Significance of Neandertal and Denisovan Genomes in Human Evolution". Annual Review of Anthropology. 42: 433–449. doi:10.1146/annurev-anthro-092412-155548.
- 1 2 Bar-Yosef, O. (1992). The role of western Asia in modern human origins. Philosophical Transactions: Biological Sciences, 337(1280), 193–200.
- ↑ Grün, R., Stringer, C., McDermott, F., Nathan, R., Porat, N., Robertson, S., … McCulloch, M. (2005). U-series and ESR analyses of bones and teeth relating to the human burials from Skhul. Journal of Human Evolution, 49(3), 316–334. http://doi.org/10.1016/j.jhevol.2005.04.006
- ↑ Frumkin, A., Bar-Yosef, O., & Schwarcz, H. P. (2011). Possible paleohydrologic effect on hominin migration and occupation of the Levantine Middle Paleolithic. Journal of Human Evolution2, 60(4), 437–451. http://doi.org/10.1016/j.jhevol.2010.03.010
- 1 2 3 4 Hallin, K. A., Schoeninger, M. J., & Schwarcz, H. P. (2012). Paleoclimate during Neandertal and anatomically modern human occupation at Amud and Qafzeh, Israel: the stable isotope data. Journal of Human Evolution, 62(1), 59–73. http://doi.org/10.1016/j.jhevol.2011.09.005
- ↑ Harrison, D. L., & Bates, P. J. J. (1991). Felidae. In The mammals of Arabia (2nd ed., pp. 156–172). Sevenoaks: Harrison Zoological Museum.
- ↑ Rampino, M. R., & Self, S. (1992). Volcanic Winter and accelerated glaciation following the Toba super-eruption. Nature, 369(6390), 50–52. http://doi.org/10.1038/359050a0
- 1 2 3 Shea, J. J. (2008). Transitions or turnovers ? Climatically-forced extinctions of Homo sapiens and Neanderthals in the East Mediterranean Levant. Quaternary Science Reviews, 1–18. http://doi.org/10.1016/j.quascirev.2008.08.015
- ↑ Tchernov, E. (1989). The Middle Paleolithic mammalian sequence and its bearing on the origin of Homo sapiens in the southern Levant. In O. Bar-Yosef & B. Vandermeersch (Eds.), Investigations in South Levantine Prehistory (pp. 25–42). Oxford: Archaeopress.
- 1 2 Belmaker, M., & Hovers, E. (2011). Ecological change and the extinction of the Levantine Neanderthals: implications from a diachronic study of micromammals from Amud Cave, Israel. Quaternary Science Reviews, 30, 3196–3209. http://doi.org/10.1016/j.quascirev.2011.08.001
- ↑ Lieberman, D. E., & Shea, J. J. (1994). Behavioral Differences between Archaic and Modern Humans in the Levantine Mousterian. American Anthropologist, 96(2), 300–332. http://doi.org/10.1525/aa.1994.96.2.02a00030
- ↑ D.O. Henry; M. Belmaker; S.M. Bergin (2017). "The effect of terrain on Neanderthal ecology in the Levant". Quaternary International. 435 (Part A): 94–105. doi:10.1016/j.quaint.2015.10.023.
- ↑ M. Şenyürek (1949). "Türk Tarih Kurumu Adına Yapılan Karain Kazısında Bulunan İki Fosil Dişe Dair Kısa Ön Rapor". Belleten (in Turkish). 52: 833–837. Cited in Aytek and Harvati (2016)
- 1 2 A.I. Aytek; K. Harvati (2016). "The Human Fossil Record from Turkey". In K. Harvati. Paleoanthropology of the Balkans and Anatolia: Human Evolution and its Context. Springer Netherlands.
- ↑ I. Yalçınkaya (1988). "9. Kazı Sonuçları Toplantısı". 1986 Yılı Karain Kazıları (in Turkish). pp. 15–37. Cited in Aytek and Harvati (2016)
- ↑ J.F. Ewing. "A probable Neanderthaloid from Ksâr 'Akil, Lebanon". American Journal of Physical Anthropology. 21. pp. 101–104.
- ↑ J.K. Williams; C.A. Bergman (2010). "Upper Paleolithic Levels XIII-VI (A and B) from the 1937-1938 and 1947-1948 Boston College Excavations and the Levantine Aurignacian at Ksar Akil, Lebanon". Paléorient. 36 (2): 117–161.
- 1 2 3 K.P. Oakley; B.G. Campbell; T.I. Molleson (1975). Catalogue of fossil hominids: Part III. Americas, Asia, Australasia. London: British Museum (Natural History).
- ↑ S. El Zaatari. "The central Levantine corridor: The Paleolithic of Lebanon". Quaternary International. in press. pp. 1–15. doi:10.1016/j.quaint.2017.06.047.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 A.-M. Tillier; B. Vandermeersch; B. Arensburg; M. Chech (2003). "New human remains from Kebara Cave (Mount Carmel). The place of the Kebara hominids in the Levantine Mousterian fossil record". Paléorient. 29 (2): 35–62.
- ↑ i1-r, i1-l, m1-r, i2-r, lower c-r and c-l, m1-r, m1-l, m2-r, M1
- 1 2 3 4 5 6 7 A. Le Cabec; P. Gunz; K. Kupczik; J. Braga; J.-J. Hublin (2013). "Anterior tooth root morphology and size in Neanderthals: Taxonomic and functional implications". Journal of Human Evolution. 64: 169–193. doi:10.1016/j.jhevol.2012.08.011.
- ↑ H. Valladas; J.L. Joron; G. Valladas; B. Arensburg; O. Bar-Yosef; et al. (1987). "Thermoluminescence dates for the Neanderthal burial site at Kebara in Israel". Nature. 330: 159–160. doi:10.1038/330159a0.
- ↑ H.P. Schwarcz; W.M. Buhay; R. Grün; H. Valladas; E. Tchernov; O. Bar-Yosef; B. Vandermeersch (1989). "ESR dating of the Neanderthal site, Kebara Cave, Israel". Journal of Archaeological Science. 16 (6): 653–659. doi:10.1016/0305-4403(89)90029-0.
- ↑ P. Smith; B. Arensburg (1977). "A Mousterian skeleton from Kebara Cave". In B. Arensburg; O. Bar-yosef. Moshe Stekelis Memorial Volume. Jerusalem: The Israel Exploration Society. pp. 164–176.
- ↑ B. Arensburg; O. Bar-Yosef; M. Chech; P. Goldberg; H. Laville; L. Meignen; Y. Rak; E. Tchernov; A.-M. Tillier; B. Vandermeersch (1985). "Une sépulture néandertalienne dans la grotte de Kébara (Israël)". Comptes Rendus de l'Académie des Sciences (in French) (300): 227–230.
- ↑ A. Keith (1931). New Discoveries Relating to the Antiquity of Man. London: William and Norgate. pp. 204–208. Retrieved 15 February 2017.
- ↑ A. Jelinek. "The Middle Paleolithic in Southern Levant, with comments on the appearance of modern Homo sapiens". In A. Ronen. The transition from Lower to Middle Paleolithic and the origin of modern man.
- ↑ R. Grün; C.B. Stringer. "Tabun revisited: Revised ER chronology and new ESR and U-series analyses of dental material from Tabun C1". Journal of Human Evolution. 39 (6): 601–612.
- ↑ B. Vandermeersch. "Neanderthal Geographical and Chronological Variation". In S. Condemi; G.-C. Weniger. Continuity and Discontinuity in the Peopling of Europe: One Hundred and Fifty Years of Neanderthal Study.
- ↑ A.-M. Tillier (2005). "The Tabun C1 skeleton: A Levantine Neanderthal?". Mitekufat Haeven: Journal of the Israel Prehistoric Society. 35: 439–450.
- 1 2 3 E. Trinkaus (1987). "The Upper Pleistocene Human Molar from Me'arat Shovakh (Mugharet Esh-Shubbabiq), Israel". Paléorient. 13 (1). pp. 95–100.
- 1 2 S. Binford (1966). "Me'arat Shovakh (Mugharet esh-Shabbabiq)". Israel Exploration Journal. 16 (2): 96–103.
- 1 2 H. Valladas; N. Mercier; L. Froget (1999). "TL dates for the Neanderthal site of the Amud Cave, Israel". Journal of Archaeological Science. 26. pp. 259–268. doi:10.1006/jasc.1998.0334.
- ↑ Suzuki, Hisashi; Takai, F. (1970). The Amud man and his cave site. Tokyo: Keigaku Publishing Co.
- ↑ Y. Rak; W.H. Kimbel; E. Hovers (1994). "A Neandertal infant from Amud Cave, Israel". Journal of Human Evolution. 26 (4).
- ↑ T. Akazawa; Y. Dodo; S. Muhesen; A. Abdul-Salam; Y. Abe (1993). "The Neanderthal Remains from Dederiyeh Cave, Syria: Interim Report". Anthropological Science. 101 (4): 361–387.
- ↑ T. Akazawa; S. Muhesen; H. Ishida; O. Kondo; C. Griggo (1999). "New Discovery of a Neanderthal Child Burial from the Dederiyeh Cave in Syria". Paléorient. 25 (2): 129–142.
- ↑ T.D. Stewart (1959). "Restoration and study of the Shanidar I Neanderthal skeleton in Baghdad, Iraq". Year Book of the American Philosophical Society for 1958: 274–278.
- ↑ T.D. Stewart (1961). "The Skull of Shanidar II". Sumer. 17: 97–106.
- ↑ R.S. Solecki (1960). "Three adult Neanderthal skeletons from Shanidar cave, northern Iraq". Annual Report of the Smithsonian Institution for 1959: 603–635.
- ↑ T.D. Stewart (1963). "Shanidar skeletons IV and VI". Sumer. 19: 8–26.
- ↑ E. Trinkaus (1977). "The Shanidar 5 Neanderthal skeleton". Sumer. 33: 35–41.
- 1 2 E. Pomeroy; M.M. Lahr; F. Crivellaro; L. Farr; T. Reynolds; C.O. Hunt; G. Barker. "Newly discovered Neanderthal remains from Shanidar Cave, Iraqi Kurdistan, and their attribution to Shanidar 5". Journal of Human Evolution. 111. pp. 102–118. doi:10.1016/j.jhevol.2017.07.001.
- ↑ M.S. Şenyürek (1957). "The skeleton of the fossil infant found in Shanidar cave, northern Iraq". Anatolia. 2: 49–55.
- ↑ M.S. Şenyürek (1957). "A further note on the paleolithic Shanidar infant". Anatolia. 2: 111–121.
- ↑ L.W. Cowgill; E. Trinkaus; M.A. Zeder (2007). "Shanidar 10: A Middle Paleolithic immature distal lower limb from Shanidar Cave, Iraqi Kurdistan". Journal of Human Evolution. 53 (2): 213–223.
- ↑ B. Wood, ed. (2013). Wiley-Blackwell Encyclopedia of Human Evolution. Wiley-Blackwell. ISBN 978-1-118-65099-8.
- ↑ E. Trinkaus; F. Biglari (2006). "Middle Paleolithic Human Remains from Bisitun Cave, Iran". Paléorient. 32 (2): 105–111.