Beta cell
| Beta cell | |
|---|---|
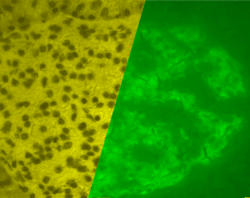 A pancreatic islet in a pig. The left image is a brightfield image created using hematoxylin stain; nuclei are dark circles and the acinar pancreatic tissue is darker than the islet tissue. The right image is the same section stained by immunofluorescence against insulin, indicating beta cells. | |
| Details | |
| Location | Pancreatic islet |
| Function | Insulin secretion |
| Identifiers | |
| Latin | endocrinocytus B; insulinocytus |
| TH | H3.04.02.0.00026 |
|
Anatomical terms of microanatomy | |
Beta cells (β cells) are a type of cell found in the pancreatic islets of the pancreas. They make up 65–80% of the cells in the islets.
Function
The primary function of a beta cell is to store and release insulin. Insulin is a hormone that brings about effects which reduce blood glucose concentration. Beta cells can respond quickly to spikes in blood glucose concentrations by secreting some of their stored insulin while simultaneously producing more.
Voltage-gated calcium channels and ATP-sensitive potassium ion channels are embedded in the cell surface membrane of beta cells. These ATP-sensitive potassium ion channels are normally open and the calcium ion channels are normally closed. Potassium ions diffuse out of the cell, down their concentration gradient, making the inside of the cell more negative with respect to the outside (as potassium ions carry a positive charge). At rest, this creates a potential difference across the cell surface membrane of -70mV.
When the glucose concentration outside the cell is high, glucose molecules move into the cell by facilitated diffusion, down its concentration gradient through the GLUT2 transporter.[1] Since beta cells use glucokinase to catalyze the first step of glycolysis, metabolism only occurs around physiological blood glucose levels and above. Metabolism of the glucose produces ATP, which increases the ATP to ADP ratio.[2]
The ATP-sensitive potassium ion channels close when this ratio rises. This means that potassium ions can no longer diffuse out of the cell.[3] As a result, the potential difference across the membrane becomes more positive (as potassium ions accumulate inside the cell). This change in potential difference opens the voltage-gated calcium channels, which allows calcium ions from outside the cell to diffuse in down their concentration gradient. When the calcium ions enter the cell, they cause vesicles containing insulin to move to, and fuse with, the cell surface membrane, releasing insulin by exocytosis.[4]
Other hormones
- C-peptide, which is secreted into the bloodstream in equimolar quantities to insulin. C-peptide helps to prevent neuropathy and other vascular deterioration related symptoms of diabetes mellitus.[5] A practitioner would measure the levels of C-peptide to obtain an estimate for the viable beta cell mass.[6]
- Amylin, also known as islet amyloid polypeptide (IAPP).[7] The function of amylin is to slow the rate of glucose entering the bloodstream. Amylin can be described as a synergistic partner to insulin, where insulin regulates long term food intake and amylin regulates short term food intake.
Clinical significance
- Type 1 diabetes mellitus, also known as insulin dependent diabetes, is believed to be caused by an autoimmune response where the body's own immune system attacks the beta cells and destroys them. This means the body can no longer produce and secrete insulin into the blood and regulate the blood glucose concentration.
- Type 2 diabetes mellitus, also known as non insulin dependent diabetes, is caused by many factors including: age; family history; obesity and consuming a diet high in simple sugars. The beta cells, however, can still secrete insulin but the body has developed a resistance and its response to insulin has declined. It is believed to be due to the decline of specific receptors on the surface of the liver and muscle cells which lose their ability to respond to insulin that circulates in the blood.[8][9]
- Insulinoma is a rare tumor derived the neoplasia of beta cells. Insulinomas are usually benign, but may be medically significant and even life-threatening due to recurrent and prolonged attacks of hypoglycemia.
Diabetes mellitus can be experimentally induced for research purposes by streptozotocin[10] or alloxan,[11] which are specifically toxic to beta cells.
Research
There is much research being conducted that attempts to artificially reconfigure beta cells in order to use them in clinical applications:
Type 1 diabetes
Type 1 Diabetes is caused by an auto-immune mediated destruction of the insulin producing beta cells in the body. The destruction of these cells reduces the body’s ability to respond to glucose levels in the body, therefore making it nearly impossible to properly regulate glucose and glucagon levels in the bloodstream. The body destroys 70-80% of beta cells, leaving only 20–30% of functioning cells. .This can cause the patient to experience hyperglycemia, which leads to other adverse short-term and long-term conditions. It has been shown that the symptoms of diabetes can be successfully controlled with methods such as regular doses of insulin and sustaining a proper diet. These methods, however, can be tedious and cumbersome to continuously perform on a daily basis. Since Type 1 diabetes is caused by damage sustained to the beta cells, the most effective strategy would be to investigate solutions to repair damaged beta cells or artificially develop and regenerate beta cells in vivo (in the body.)
Research has shown that beta cells can be differentiated from human pancreas progenitor cells. These differentiated beta cells, however, often lack much of the structure and markers that beta cells need to perform their necessary functions. Examples of the anomalies that arise from beta cells differentiated from progenitor cells include a failure to react to environments with high glucose concentrations, an inability to produce necessary beta cell markers, and abnormal expression of glucagon along with insulin.
In order to successfully re-create functional insulin producing beta cells, studies have shown that manipulating cell-signal pathways in early stem cell development will lead to those stem cells differentiating into viable beta cells. Two key signal pathways have been shown to play a vital role in the differentiation of stem cells into beta cells: the BMP4 pathway and the kinase C. Targeted manipulation of these two pathways has shown that it is possible to induce beta cell differentiation from stem cells. These variations of artificial beta cells have shown greater levels of success in replicating the functionality of natural beta cells, although the replication has not been perfectly re-created yet.
Many studies have shown that it is possible to regenerate beta cells in vivo in some animal models. Research in mice studies have shown that beta cells can often regenerate to the original quantity number after the beta cells have undergone some sort of stress test, such as the intentional destruction of the beta cells in the mice subject or once the auto-immune response has concluded. While these studies have conclusive results in mice, beta cells in human subjects may not possess this same level of versatility. Investigation of beta cells following acute onset of Type 1 diabetes has shown little to no proliferation of newly synthesized beta cells, which suggests that the results seen in the mice models might not occur in human subjects.
It appears that much work has to be done in the field of regenerating beta cells.[12] Just as in the discovery of creating insulin through the use of recombinant DNA, the ability to artificially create stem cells that would differentiate into beta cells would prove to be an invaluable resource to patients suffering from Type 1 diabetes. An unlimited amount of beta cells produced artificially would provide therapy to many of the patients who are affected by Type 1 diabetes.
In 2018, a new drug was developed in Britain that may be able to regenerate beta cells.[13]
See also
References
- ↑ De Vos, Anick; Heimberg, Harry; Quartier, Erik; Huypens, Peter; Bouwens, Luc; Pipeleers, Daniel; Schuit, Frans (1995). "Human and rat beta cells differ in glucose transporter but not in glucokinase gene expression". Journal of Clinical Investigation. 96 (November): 2489–95. doi:10.1172/JCI118308. PMC 185903.
- ↑ Santulli, Gaetano; Pagano, Gennaro; Sardu, Celestino; Xie, Wenjun; Reiken, Steven; D’Ascia, Salvatore Luca; Cannone, Michele; Marziliano, Nicola; Trimarco, Bruno; Guise, Theresa A.; Lacampagne, Alain; Marks, Andrew R. (2015). "Calcium release channel RyR2 regulates insulin release and glucose homeostasis". Journal of Clinical Investigation. 125 (5): 1968–78. doi:10.1172/JCI79273. ISSN 0021-9738. PMC 4463204. PMID 25844899.
- ↑ Keizer J, Magnus G (1989). "ATP-sensitive potassium channel and bursting in the pancreatic beta cell. A theoretical study". Biophysical Journal. 56 (2): 229–42. Bibcode:1989BpJ....56..229K. doi:10.1016/S0006-3495(89)82669-4. PMC 1280472. PMID 2673420.
- ↑ Lang V, Light PE (2010). "The molecular mechanisms and pharmacotherapy of ATP-sensitive potassium channel gene mutations underlying neonatal diabetes". Pharmgenomics. Pers. Med. 3: 145–61. doi:10.2147/PGPM.S6969. PMC 3513215. PMID 23226049.
- ↑ Ido Y; Vindigni A; Chang K; Stramm L; Chance R; Heath WF; et al. (1997). "Prevention of vascular and neural dysfunction in diabetic rats by C-peptide". Science. 277 (5325): 563–66. doi:10.1126/science.277.5325.563. PMID 9228006.
- ↑ Hoogwerf B, Goetz F (1983). "Urinary C-peptide: a simple measure of integrated insulin production with emphasis on the effects of body size, diet, and corticosteroids". J Clin Endocrinol Metab. 56 (1): 60–67. doi:10.1210/jcem-56-1-60. PMID 6336620.
- ↑ Moore C, Cooper G (1991). "Co-secretion of amylin and insulin from cultured islet beta-cells: modulation by nutrient secretagogues, islet hormones and hypoglycemic agents". Biochem Biophys Res Commun. 179 (1): 1–9. doi:10.1016/0006-291X(91)91325-7. PMID 1679326.
- ↑ "U.K. prospective diabetes study 16. Overview of 6 years' therapy of type II diabetes: a progressive disease. U.K. Prospective Diabetes Study Group". Diabetes Journal. Retrieved 2014-04-21.
- ↑ Rudenski A, Matthews D, Levy J, Turner R (1991). "Understanding "insulin resistance": both glucose resistance and insulin resistance are required to model human diabetes". Metabolism. 40 (9): 908–17. doi:10.1016/0026-0495(91)90065-5. PMID 1895955.
- ↑ Wang Z, Gleichmann H (1998). "GLUT2 in pancreatic islets: crucial target molecule in diabetes induced with multiple low doses of streptozotocin in mice". Diabetes. 47 (1): 50–56. doi:10.2337/diabetes.47.1.50. PMID 9421374.
- ↑ Danilova IG, Sarapultsev PA, Medvedeva SU, Gette IF, Bulavintceva TS, Sarapultsev AP (2014). "Morphological Restructuring of Myocardium During the Early Phase of Experimental Diabetes Mellitus". Anat. Rec. 298 (2): 396–407. doi:10.1002/ar.23052.
- ↑ Mahla RS (2016). "Stem cells application in regenerative medicine and disease threpeutics". International Journal of Cell Biology. 2016 (7): 19. doi:10.1155/2016/6940283. PMID 27516776.
- ↑ "'Ground-breaking' diabetes insulin drug trialled in Cardiff - BBC News".