Langetermijngeheugen
In het langetermijngeheugen wordt informatie voor lange tijd in de hersenen opgeslagen. Het wordt doorgaans in twee ondervormen onderverdeeld: het declaratieve en het niet-declaratieve geheugen. In plaats hiervan worden ook de termen expliciet (bewust) en impliciet (onbewust) wel gebruikt.
.jpg)
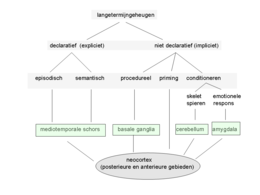
Het declaratieve geheugen bevat kennis omtrent bepaalde gebeurtenissen en feiten die zich in ons leven hebben voorgedaan, en die ook toegankelijk zijn voor het bewustzijn. Daarentegen omvat het niet-declaratieve geheugen verschillende ondervormen van impliciet geheugen zoals het procedurele geheugen, perceptuele priming en gedrag dat via associatieve leerprocessen, zoals conditioneren, tot stand is gekomen.
Geheugen en neurale netwerken
Indrukken of informatie uit de buitenwereld worden door leerprocessen in het geheugen opgeslagen. Deze opslag vindt plaats in netwerken van zenuwcellen via een proces van consolidatie. Een belangrijke rol hierbij spelen de synapsen (verbindingen tussen zenuwcellen). Volgens de theorie van Donald Hebb wordt namelijk een synaps versterkt als zowel het pre- als postsynaptische neuron tegelijk actief zijn. Versterking van deze verbindingen heet potentiëring. Veel theorieën betreffende het leren beschouwen langetermijnpotentiëring (LTP) en het tegengestelde proces langetermijndepressie als de cellulaire basis van leren en geheugen. LTP werd in 1966 ontdekt door Terje Lømo en is sindsdien een populair onderwerp in het neurowetenschappelijk onderzoek. Een verzameling van zenuwcellen waar bepaalde informatie is opgeslagen (bijvoorbeeld het beeld van een object zoals een bal of een huis, het gezicht van grootmoeder of van een aangeleerde handeling) wordt in de cognitieve psychologie en neurobiologie representatie genoemd. Vermoedelijk zijn hierbij grotere netwerken of ensembles van zenuwcellen betrokken.
Neurale basis van het declaratieve geheugen
De indeling van het langetermijngeheugen is mede gebaseerd op hersenonderzoek bij mensen en dieren. Een belangrijk ontdekking daarbij was dat het declaratieve geheugen wordt gevormd door leerprocessen waarbij vooral de mediale temporale schors (inclusief de hippocampus) is betrokken. Beschadiging van de hippocampus of de aangrenzende gebieden in de mediale temporale schors, leidt vaak tot ernstige stoornissen in het vastleggen van nieuwe kennis van feiten en gebeurtenissen.
- Patiënt H.M.
Een ‘beroemd’ geval waarbij deze stoornissen optraden was Brenda Milners patiënt H.M., die na operatieve verwijdering van delen van de hippocampus en omliggende delen zoals de gyrus parahippocampalis geen nieuwe feiten of gebeurtenissen meer voor lange tijd in zijn geheugen kon opslaan. Daarentegen was zijn kortetermijngeheugen (bijvoorbeeld het voor korte tijd kunnen onthouden van telefoonnummers) nagenoeg intact. Ook bleek H.M. in staat tot het leren van perceptueel-motorische vaardigheden. De geheugenstoornis die zich bij H.M. voordeed, wordt anterograde amnesie (geheugenverlies) genoemd.
- Dieronderzoek
Ook onderzoek met andere patiënten, en onderzoek met apen van Squire[1] toonde later het belang aan van de mediale temporale schors voor de vorming van het declaratieve langetermijngeheugen. Squire e.a. maakten daarbij gebruik van een ingenieuze taak, de zogeheten delayed nonmatching to sample-taak die door Mishkin was bedacht. In deze taak krijgt het proefdier bij elke leertrial twee plateaus te zien. Het ene plateau is gemarkeerd met een figuur die ook op de vorige trial te zien en beloond was (bijvoorbeeld een kruis), en het andere plateau is gemarkeerd met een niet eerder vertoonde figuur (bijvoorbeeld een cirkel). Het voedsel wordt nu steeds onder de nieuwe figuur geplaatst. Bij laesies van de hippocampus van de apen bleek in deze taak het leren kiezen van nieuwe objecten aanzienlijk bemoeilijkt.
- Waar zit het geheugen?
De structuren van de mediale temporale schors vormen overigens niet zelf de plaats waar het langetermijngeheugen zich nestelt. Dit gebeurt vermoedelijk in hogere gebieden van het achterdeel van de cortex cerebri en de temporale schors. Met het consolidatieproces kan een relatief lange tijd gemoeid zijn. Dit verklaart waarom bij laesies in het mediale temporale gebied (zoals ook bij patiënt H.M.) vaak een retrograde amnesie van enkele maanden of nog langer optreedt. Kennelijk is dan de consolidatie van oude kennis (=gebeurtenissen die zich voor het moment van de laesie voordeden) nog niet voltooid.
- Twee vormen: episodisch en semantisch geheugen
Het declaratieve geheugen wordt ten slotte nog verder onderverdeeld in het episodische geheugen (ons persoonlijk gekleurd biografisch geheugen voor gebeurtenissen die in het verleden hebben gespeeld) en het semantische geheugen (onze kennis van de wereld: zoals kennis van de taal, weten hoe onze koning heet, en hoe je een ei bakt). Het is echter niet waarschijnlijk dat het episodische en semantische geheugen ook kwalitatief verschillende systemen in de hersenen representeren. Het enige verschil lijkt te zijn dat het semantische geheugen hechter verankerd is in netwerken van de posterieure hersenen. Dit heeft mogelijk weer te maken met het feit dat in het dagelijks leven semantische kennis of informatie veel intensiever wordt gebruikt dan episodische kennis of informatie. Ook nestelen trauma's zich in het episodische geheugen.
Neurale basis van het niet-declaratieve geheugen
Het niet-declaratieve geheugen vormt een heterogene categorie. Dit slaat niet alleen op de aard van de betrokken geheugensystemen, maar ook op de gebieden in de hersenen die hiermee verbonden zijn.
- Procedureel geheugen. Hieronder worden motorische en cognitieve vaardigheden (Engels: skills) verstaan. Bij de vorming van het procedurele geheugen spelen subcorticale gebieden zoals de basale kernen een rol. Bij de permanente opslag in de hersenen lijken vooral de motorische schorsgebieden betrokken te zijn.
- Priming slaat op verwerving van perceptuele kennis, als gevolg van eerdere blootstelling aan een prikkel. Een voorbeeld is de lexicale decisietaak waarin een proefpersoon moet aangeven of een woord een echt woord (HOND) of onzinwoord is (DNOK). Dit gaat doorgaans sneller als het echte woord in een eerder aangeboden lijst voorkwam. Het is net alsof door het eerder aangeboden woord een gebiedje in de hersenen al is ’klaargezet’. Primingeffecten blijken een gevolg te zijn van activatie van elementaire perceptuele eenheden, zogenaamde modulen, in de posterieure (vooral sensorische) schorsgebieden.
- Conditioneren. Hierbij kan een onderscheid worden gemaakt tussen klassiek conditioneren (associatie van een voorwaardelijke prikkel zoals een toon, met een onvoorwaardelijke prikkel zoals een hard geluid, elektrische schok of luchtstootje tegen het hoornvlies van het oog), en operante conditionering (hierbij wordt gedrag gevolgd door beloning of straf). Het cerebellum speelt vooral een rol bij klassiek conditioneren van motorisch gedrag. Ook emotioneel gedrag (zoals vrees- en schrikreacties) blijkt vaak te berusten op deze meer elementaire vormen van associatief leren. Een belangrijke structuur vormen hier de amygdalae (amandelkernen). Deze zijn vooral betrokken bij aangeleerde vreesreacties.
Korte- en langetermijngeheugen: gescheiden of identieke systemen in de hersenen?
In de cognitieve psychologie worden het korte- en het langetermijngeheugen vaak gezien als gescheiden systemen. Dit onderscheid is vooral functioneel van aard. Het is namelijk vooral bepaald door de duur van de opslag van informatie in de hersenen: iets wordt voor korte tijd of langere tijd in het geheugen vastgehouden of bewust beleefd. Neurobiologen zoals Joachim Fuster zijn van mening dat er geen bewijs is dat het korte- en langetermijngeheugen ook met aparte neurale netwerken corresponderen.[2] Dit geldt vooral voor de plaats van opslag van informatie in de hersenen. Het kortetermijngeheugen kan namelijk worden gezien als een tijdelijke activering van dezelfde neurale netwerken die ook het langetermijngeheugen vormen. Een herinnering aan een vakantie is bijvoorbeeld in passieve of sluimerende toestand onderdeel van het langtermijngeheugen. Zodra wij echter deze herinnering weer 'voor de geest halen', verkeren de neuronen van hetzelfde netwerk voor korte tijd in een actieve toestand.
Rol van de prefrontale cortex
De traditionele opvatting is dat de prefrontale cortex vooral een sturende/regelende functie heeft. Verschillende delen van de prefrontale cortex hebben lange verbindingsvezels met specifieke delen in de meer naar achter gelegen delen van de hersenen. De prefrontale cortex is vermoedelijk niet zélf de plaats waar informatie in het langtermijngeheugen wordt opgeslagen. Wél lijkt het gebied een actieve rol te spelen bij de codering (inprenting) en terugvinden (retrieval) van informatie die in andere delen van de hersenen (o.a. de temporale schors) is opgeslagen. Met behulp van beeldvormende technieken hebben o.a. Tulving[3] aannemelijk gemaakt dat linker- en rechterdelen van de prefrontale cortex betrokken zijn bij respectievelijk de codering en het terugvinden van informatie in het episodische geheugen. Vooral het rechterdeel van de dorsolaterale prefrontale cortex lijkt een belangrijke rol te spelen bij het terugvinden van informatie[4]
Zie ook
Bronnen, noten en/of referenties
|