Ala (rondworm)
Ala (meervoud alae) zijn een of twee, vleugelachtige verdikkingen aan de buitenkant van een rondworm en worden gevormd door het onderhuids weefsel (hypodermaal weefsel). De alae worden gevormd door een oxidatief proces waarbij peroxidase inwerkt op de eiwitgebonden tyrosine-residuen. Hierbij ontstaan plooien door het radiaal krimpen van de door de laterale onderhuidse weefselcellen ('naadcellen') uitgescheiden eiwitten. De laterale onderhuidse weefselcellen zijn gerangschikt in twee in de lengte liggende rijen van 16 cellen aan de linker- en rechterkant van het lichaam en ingebed in het cilindrische hypodermale 7-syncytium.[1]
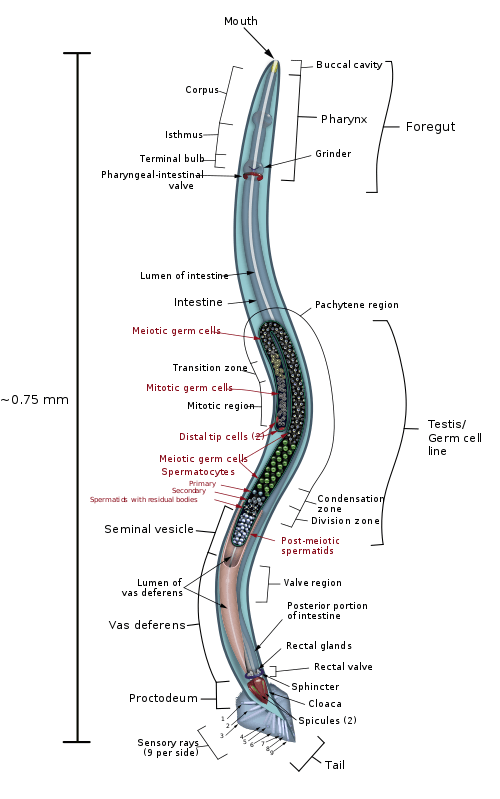
De alae komen vooral voor bij de dauerlarve, maar kunnen ook bij de volwassen rondworm voorkomen.
Plaats
- Caudale alae of bursae zitten bij de staart van de mannelijke rondworm en hebben sensorische stralen.
- Cervicale alae zitten bij de kop van de rondworm en zijn alleen bekend in de parasitaire orden Strongylida, Ascaridida, Spirurida en bij Benthimermithida bij het geslacht Trophomera.
- Cervicale laterale alae die voorkomen bij mannetjes van Trophomera-soorten.[2]
- Alae preamfidiaal, zich uitstrekkend over de gehele afstand van de voorkant van de kop tot aan de amfide;
- Alae preafaidiaal, zich slechts uitstrekkend over een deel van de afstand van de voorkant van de kop tot aan de amfide;
- Alae, die zich uitstrekken tot preamfidiale en postamfidiale delen.
- Cervicale laterale alae die voorkomen bij mannetjes van Trophomera-soorten.[2]
- Laterale alae zitten aan de buitenkant van het lichaam van de rondworm.[3]
- Longitudinale alae verlengen het lichaam en zitten in het algemeen lateraal of sublateraal van het lichaam van de rondworm.
 Cervicale alae bij de aarsmade
Cervicale alae bij de aarsmade_FIGS_13-20.tif.jpg) 18: Bursa met sensorische stralen bij Torrestrongylus tetradorsalis
18: Bursa met sensorische stralen bij Torrestrongylus tetradorsalis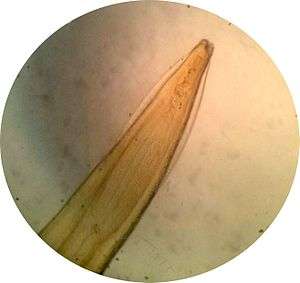 Cervicale alae bij de hondenspoelworm (Toxocara canis)
Cervicale alae bij de hondenspoelworm (Toxocara canis) Cervicale alae bij de kattenspoelworm (Toxocara cati)
Cervicale alae bij de kattenspoelworm (Toxocara cati)
Functie
De functie van de alae is nog niet duidelijk. Ze zouden kunnen bijdragen aan de versterking van de cuticula, de beweging van de rondworm of opslag van lipiden. Het grootste gedeelte van de structuur van de alae van Caenorhabditis elegans bevatten echter de ZP-domein eiwitten (CUT-1, CUT-3, CUT-5 CUT=cuticula). Het ZP-domein is betrokken bij de vorming van de matrixlaag en heeft vermoedelijk ook functies bij het waarnemen van feromonen en het waarnemen van geuren (olfactorische signaaltransductie).[4][5] Ondanks het structurele karakter van het ZP-domein eiwit is het niet het ideale eiwit, dat alleen voor de sterkte kan zorgen. Daar waar ZP-domein eiwitten worden gevonden, worden deze gevonden in een vaste of schijnbare associatie met signaaltransductie door interactie met een externe of vijandige omgeving.
De alae lijken receptoren van zenuwcellen te zijn, die reageren op slechts een handvol verschillende moleculen specifiek voor elke rondwormensoort. Er kunnen op een bepaald moment tot een half dozijn soorten receptoren op de alae aanwezig zijn en van elk type kunnen er dan veel tegelijk voorkomen. De concentratie doelmoleculen in de omgeving van de rondworm, die de receptor stimuleren, kunnen vervolgens door de rondworm worden gemeten en als er een drempelwaarde wordt bereikt, zal er tot actie worden overgegaan. Deze acties kunnen bestaan uit het zorgen van toegang tot de dauerlarve, het verlaten van het dauerlarvestadium (door feromonen en of de aanwezigheid van een indicator voor voedselbeschikbaarheid), voor geslachtelijke voortplanting waar het gebied rond de vulva en gonaden van het vrouwtje en het mannetje de familie van RAM-5-receptoren gebruiken voor het waarnemen van contact tussen de voortplantingsorganen.[6]
{kind=link}