Actiepotentiaal
In de fysiologie is een actiepotentiaal een golf van elektrische ontlading over de membraan van een exciteerbare, dus prikkelbare cel, zoals een neuron of een spiercel.[1] Actiepotentialen vormen een essentiële eigenschap van dierlijk leven, maar komen ook voor in sommige planten. Ze maken het mogelijk om snel informatie te verzenden tussen verschillende weefsels. Het zenuwstelsel maakt uitvoerig gebruik van actiepotentialen, om informatie tussen zenuwcellen onderling uit te wisselen, maar ook tussen zenuwcellen en andere celtypen, zoals spieren of klieren, of tussen spiercellen onderling, zoals in de hartspier.[2]
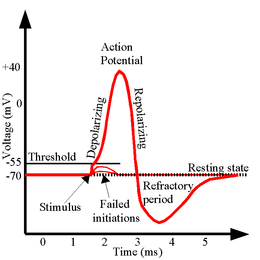
Mechanisme
Om een actiepotentiaal te genereren, zijn er verschillende factoren nodig. Ten eerste moet er een extracellulaire prikkel zijn, bijvoorbeeld een neurotransmitter. Dit leidt tot een depolarisatie van de celmembraan, die leidt tot het verplaatsen van deze depolarisatie langs het axon in een zichzelf ondersteunend proces.
Membraanpotentiaal
De membraanpotentiaal is de elektrische spanning die staat over de membraan van een cel. De potentiaal ontstaat door verschillende concentratiegradiënten van positieve en negatieve ionen (= elektrische lading!) aan weerszijden van de membraan: aan de buitenkant (de extracellulaire zijde) van de membraan zijn er meer positieve ionen (vooral natriumionen) dan aan de binnenkant (het cytoplasma, waarin vooral kaliumionen) van de cel.
Fysiologie
Door het binden van de transmitter worden de kanalen geopend waardoor de membraanpotentiaal verandert. In rust is deze ongeveer -70 millivolt (mV) ten opzichte van het externe milieu. Door het binden verandert de geleidbaarheid voor een ion (meestal natrium) die daardoor de cel instroomt. Door deze instroom stijgt de membraanpotentiaal naar een bepaalde grenswaarde van ongeveer -50 mV. Als deze grenswaarde (in het Engels: threshold) bereikt wordt, begint een proces van zichzelf versterkende activatie. Dit komt doordat er spanningsgevoelige kanalen in de celmembraan zitten die opengaan boven deze waarde. Hierdoor stijgt de membraanpotentiaal nog sterker tot een waarde van ongeveer +25 mV: de cel is nu gedepolariseerd. Als dit de enige kanalen zouden zijn dan zou de gedepolariseerde toestand eindeloos gehandhaafd blijven. Om weer te repolariseren gaat een tweede soort kanalen open, namelijk de kaliumkanalen. Dit leidt tot de uitstroom van kaliumionen en een daling van de membraanpotentiaal. Zodra de potentiaal weer onder de grenswaarde zit gaan de kalium kanalen weer langzaam dicht, maar doordat deze kanalen iets langer open blijven staan wordt de cel te sterk gepolariseerd (ook wel hyperpolarisatie genoemd). Doordat spanningsafhankelijke natriumkanalen inactiveren aan het einde van een actiepotentiaal ontstaat een refractaire periode, waarin een cel niet in staat is om een actiepotentiaal te genereren.
Model voor actiepotentialen
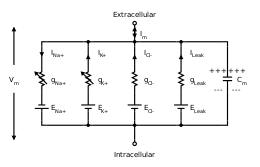
Alan Hodgkin en Andrew Huxley hebben in 1952 een wiskundig model voorgesteld dat de generatie van een actiepotentiaal in een neuron beschrijft aan de hand van de geleidbaarheid van natrium- en kalium-ionkanalen.[3] Op grond van een stel differentiaalvergelijkingen kan het gedrag van een cel op stimulatie uit de omgeving voorspeld worden. Aan de hand van waarden gemeten in vivo, konden de waarden van verschillende variabelen ingevuld worden.
De eerste vergelijking is voor de n-poort (een afsluitbaar element dat snel open- en dichtgaat) van natriumkanalen, de tweede voor de m-poort (een langzaam afsluitend element) van natriumkanalen en ten slotte de vergelijking van de kaliumkanalen. Robert Fitzhugh en J. Nagumo hebben het aantal variabelen teruggebracht tot twee vergelijkingen met dezelfde resultaten door het gedrag van de n-poort en de m-poort te combineren in één vergelijking.
Mede door het gebruik van deze wiskundige modellen is de kennis van de ionkanalen in verschillende typen exciteerbare cellen enorm toegenomen. Honderden verschillende membraanmodellen zijn inmiddels ontwikkeld, voor vele verschillende celtypen, en deze modellen beschrijven meestal tientallen verschillende ionkanalen.
Het concept van de "poorten" dat door Hodgkin en Huxley is ingevoerd is zeer succesvol gebleken. Toch waren deze onderzoekers voorzichtig genoeg om niet te veronderstellen dat er daadwerkelijk kleine poortjes aanwezig zijn in de ionkanalen. Tegenwoordig wordt gedacht dat de variabelen die de status (open of dicht) van de poorten beschrijven, samen de conformatietoestand beschrijven van de moleculen waar het ionkanaal uit bestaat. In sommige conformatietoestanden laten deze moleculen de ionen passeren; in andere niet. Het lijkt er op dat deze moleculen soms tientallen of honderden relevante configuratietoestanden hebben. In sommige gevallen kan het gedrag van deze moleculen niet goed beschreven worden met "poorten", en wordt een algemenere vorm, de zogeheten markovemodellen, gebruikt.
Neurotransmitters
Als een neurotransmitter (in de spier acetylcholine, ACh) uit de presynaptische blaasjes wordt vrijgemaakt op basis van een actiepotentiaal vanuit de aanvoerende zenuw in de synapsspleet, zal deze door diffusie naar het postsynaptisch element gaan en zich daar binden aan de postsynaptische receptor. Er zijn verschillende soorten transmitters met verschillende effecten op de postsynaptische cel. Voorbeelden zijn de excitatoire transmitters zoals glutamaat, acetylcholine en serotonine. Deze moleculen kunnen zich binden aan specifieke receptoren die een kanaal vormen door de celmembraan, die semipermeabel zijn voor ionen. Ook zijn er de inhibitoire transmitters zoals gamma-aminoboterzuur (GABA) die een tegenovergesteld effect hebben op de membraanpotentiaal: zij hyperpolariseren de cel waardoor deze meer activatie vereist om alsnog te vuren.
Literatuur
- R. Fitzhugh, Impulses and physiological states in theoretical models of nerve membranes, 1182-Biophys. J., 1 (1961), p. 445-466.
- J. Nagumo, S. Arimoto, and S. Yoshizawa, An active pulse transmission line simulating 1214-nerve axons, Proc. IRL, 50 (1960), p. 2061-2070.
Referenties
|
| Zie de categorie Action potentials van Wikimedia Commons voor mediabestanden over dit onderwerp. |