Achillobator
Achillobator (/əˌkɪləˈbaɪtɔːr/ ə-KIL-ə-BAY-tor; meaning '"Achilles hero") is a genus of large dromaeosaurid theropod dinosaur that lived in Asia during the Late Cretaceous period. It was a heavy-built, ground-dwelling, bipedal carnivore that would have been an active predator, hunting with the enlarged, sickle-shaped claw on the second toe. It has an estimated length about 5 to 6 m (16 to 20 ft). The genus is monotypic, including only the type species, A. giganticus; since its first discovery and description, no more specimens have been found. The most characteristic feature of Achillobator is its primitive pelvis, differing from the rest of dromaeosaurids, also, most of its preserved elements show prominent sturdiness, a very unusual trait in dromaeosaurids. It represents the first, and largest, dromaeosaurid taxon known from the Bayan Shireh Formation.
| Achillobator | |
|---|---|
 | |
| Skeletal diagram showing preserved elements from the holotype MNUFR-15 | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Clade: | Dinosauria |
| Clade: | Saurischia |
| Clade: | Theropoda |
| Family: | †Dromaeosauridae |
| Clade: | †Eudromaeosauria |
| Subfamily: | †Dromaeosaurinae |
| Genus: | †Achillobator Perle et al. 1999 |
| Type species | |
| †Achillobator giganticus Perle et al. 1999 | |
Discovery and naming
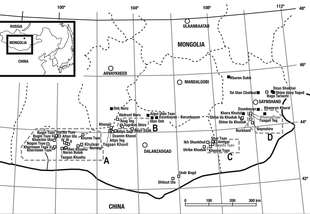
In 1989, during a field exploration conducted by the Mongolian and Russian Paleontological Expedition in the Gobi Desert, examining the outcrops at Khongil, South Central Mongolia, many dinosaur fossil discoveries were made. About 5.6 km away from the Khongil locality, a large, mostly disarticulated, partial theropod skeleton was discovered in fine-grained, medium sandstone/gray mudstone that was deposited dating back to the Late Cretaceous epoch at the Burkhant locality, Bayan Shireh Formation. The preserved specimen was found in association with a maxilla and numerous bones from the body, most notably the pelvic region. It was collected and exceptionally prepared by the assistant paleontologist Namsarai Batulseen, and labelled under the number MNUFR-15. Ten years later, in 1999, the unusual specimen was formally described and became the holotype for the new genus and species: Achillobator giganticus; the description was performed by the Mongolian paleontologist Altangerel Perle, and North American paleontologists Mark Norell and Jim Clark.[1]
However, the description was published in a very preliminary format, being not complete at all, having a few issues with the preserved elements and some bad redaction. For instance, metatarsals II and III (of unknown position) were claimed as preserved elements, but in further pages there's no mention of them, a similar case can apply to the preserved left metatarsals III and IV, which at some point are attributed to the left pes, but afterwards to the right one.[1] The paper was likely published without the knowledge of the two latter paleontologists, as indicated by a draft left in Mongolia in 1997.[2] In terms of etymology, the generic name, Achillobator, is derived from the Latin word Achillis (genitive singular of Achilles) in reference to the large Achilles tendon that supported the second pedal ungual (commonly known as "sickle" or "killer" claws) of most dromaeosaurids, and the old Mongolian word баатар (baatar, meaning hero). The specific name, giganticus, is derived from the Ancient Greek word γιγαντικός (gigantikós, meaning gigantic) in reference to the large size of the holotype, which apparently exceeds most dromaeosaurids.[1]
Chimera hypothesis
The pelvis of Achillobator features plesiomorphic saurischian characteristics compared to other dromaeosaurids. For instance, the pubis is aligned vertically and has a relatively large pubic boot (a wide expansion at the end), unlike most other dromaeosaurids, where there is generally a much smaller boot. The preserved vertebrae are very robust and features a series of pleurocoels.[1] The above differences and led Burnham et al. 2000 to suggest that Achillobator in fact, represents a paleontological chimera, arguing that only the pedal unguals may have come from a dromaeosaurid-grade dinosaur.[3] However, the specimen was actually found in semiarticulation; the assignment of the remains to a single individual is pretty much supported.[4][2] Although Achillobator features unusual and basal characteristics compared to other relatives, it is still being supported as a dromaeosaurid.[5][6]
Description

The holotype remains are fragmentary, consisting of: left maxilla with nine teeth and two alveoli; four cervical vertebrae, three dorsal vertebrae and seven caudal vertebrae; a nearly complete pelvic girdle compromising both pubis, right illium and right ischium; both femur and left tibia; left metatarsals III and IV; manual and pedal phalanges and unguals; right scapula and coracoid; isolated radius; two ribs and caudal chevrons. Traces of tail tendons were found too, but mostly isolated.[1]

The holotype was found lacking traces of feather integument, however, there are strong evidence coming from other relatives that suggest the likely presence of plumage on Achillobator, such as the smaller, and also Asiatic Velociraptor and Zhenyuanlong, and the giant North American Dakotaraptor.[7][8][9] It was among the largest dromaeosaurids; the holotype and only known individual of Achillobator was originally estimated to be three times as big as Deinonychus.[1] It has many other estimates: during the description of Mahakala in 2007, by using body mass equations to estimate size and length in many paravians, Achillobator was estimated at 4.85 m (15.9 ft) long with a weight of 347.9 kg (767 lb).[10] Novas et al. 2008, gave a length of 4.45 m (14.6 ft) and a weight of 264 kg (582 lb).[11] Turner et al. 2012 estimated its weight under 300 kg (660 lb). Also, they noted that this genus represents the second largest dromaeosaurid taxon with a tibial length of 49 cm (490 mm).[2] There are larger estimates for Achillobator: Thomas R. Holtz and Gregory S. Paul have estimated its length at 5 to 6 m (16 to 20 ft), Paul gave a weight of 250 kg (550 lb) and Holtz estimated it around 91 to 227 kg (201 to 500 lb).[12][13]
The preserved maxilla is partially complete, missing pretty much of its nasal process. It measures 29.08 cm (290.8 mm) in length, the lateral side is smooth compared to the dorsal areas, although its structure is very robust. It preserves 11 alveoli, of which 9 are filled with well-preserved teeth. The teeth are serrated and recurved with the posterior serrations being slightly larger than the anterior serrations. Perle and colleagues suggested that Achillobator had a relatively large skull, competing to those of Carnosaurs, based on the large maxilla.[1]
Postcranial skeleton
Vertebrae

Well-preserved material is known from the vertebral area, compromising very robust and somewhat large cervical, dorsal and caudal vertebrae. There are about four cervicals preserved; as stated by the authors, the anterior series of cervical vertebrae are sharply-angled developed, with the anterior articulation facet of the centrum being over the posterior facet. These characteristics are indicators of a S-shaped neck, as seen in most maniraptorans. Three dorsals were preserved with apparently two dorsal ribs. The dorsals are stocky and have pleurocoels at the lateral surfaces; some have two. Their centrum articulation facets are rounded and concave in shape. In general, the neural processes of the dorsal vertebrae show resemblance to those large ratites, such as the extinct Moa, and extant Emu, Ostrich and Cassowary, they feature protruding interspinous ligaments scars that indicates a robust and similar back to those birds. Seven caudals were found and four are successive; some traces of tail tendons were found isolated. One of the caudals belong to the transitional series (anteriormost). In all caudals the centrum is amphiplatian with the posterior articulation facet being more concave than the anterior one. As indicated by the transitional caudal, the anterior neural spines are more elongated; they disappear on the posterior caudals.[1]
Pelvic girdle and hindlimbs
The right and left femur were nicely preserved. In general terms, its femur is highly robust and longer than the tibia, a rare trait in dromaeosaurids, measuring 50.5 cm (505 mm) in length. The femora preserves detailed information of muscle attachments: on an anteromedially surface, there's a large, sculptured, concave surface that likely worked as the instertion for the muscle iliofemoralis internus in life, and the lesser trochanter preserves the insertion for the m. pubo ischio femoralis on the anterodistal margin. On the proximolateral surface of the femur, ventral to the greater trochanter, there is a tubercle that formed the attachment for the m. iliofemoralis. Insertions for m. ilio trochantericus and ischio trochantericus are located on the posterolateral surface at the proximal end of the femur. Medially, on the proximal third part of the shaft, a coarse, concave area is preserved; likely an attachment for m. pubo ischio femoralis externus-3. Below this area, a small, convex tubercle attended the insertion for m. pubo ischio femoralis-2.[1]
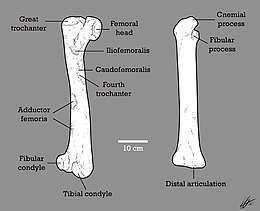
The left tibia is a little bit less robust than the femur, but more straight; it measures 49 cm (490 mm) in length. The surface of the shaft center is smooth, without traces of muscles. On the anterodorsal surface of the cnemial crest however, there is a coarse and microsculptured area that worked as the attachment for m. ilio tibialis and ambiens. Additional tissue traces in the preserved tibia are located on the proximal postero lateral surface. Here, a depression with a somewhat coarse surface likely attached prominent aponeurosis of the connected ligaments to the tibia and fibular head when the animal was alive. Apparently no other tissue surfaces are preserved.[1]
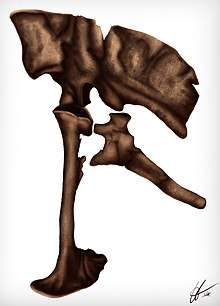
The most emblematic element from the holotype is the striking pelvis, being virtually complete; it preserves the right illium, ischium and both pubis. Although complete, the remains are slightly crushed and deformed. The illium is prominently tall with a dorsoventrally expanded preacetabular, measuring 51.3 cm (513 mm) in length and 28.8 cm (288 mm) in height. It is full of tissue traces: in a medial view, both m. caudofemoralis are preserved. On the anterior edge there are microstriations that likely gave form to the m. iliofemoralis, posteriorly to this area, traces of the m. ilio tibialis-2 are also preserved. The pubis measures 54.8 cm (548 mm) long and is very straight having a large distal pubic boot; striking resemblance to Carnosaurs. For instance, other dromaeosaurids have a very opisthopubic pelvic configuration, Achillobator however, has a primitive, propubic configuration. The large pubic boot preserves very coarse areas on the dorsal surface, indicators for the m. pubo ischio femoralis internus ventralis attachment. Its lateral view is very flat and sculptured, having numerous microstriations that probably originated the m. pubo ischio femoralis externus. The preserved ischium is shorter than the latter elements, measuring 37.8 cm (378 mm) in length. A large ridge is formed on its shaft and expands to the anterior edge, likely forming the m. adductor femoris in life. This ridge-like structure is heavy-built compared to other dromaeosaurids. On the distal lateral end, the surface is very rough, possibly attaching m. flexor tibialis internus-1 in life[1], as seen in crocodiles and other theropods.[14][15]
Distinguishing anatomical features
Following the original description, Perle and colleagues established several unique features for Achillobator that differ from other dromaeosaurids: elongated forelimbs, based on preserved radius, short pes with medium length; penultimate and ungual phalanges of the second digit robust, particularly the proximoventral process of the penultimate phalanx; the metacarpal III is irregularly elongated with a gracile end: large semicircular-oval antorbital fenestra placed vertically, the second and third accessory fenestrae are placed subvertically; maxillary teeth with anterior and posterior serrated margins; posterior denticles bigger than anterior ones; short cervical vertebrae with massive and sharp angles; elongated and flat caudal vertebrae; rodlike prezygapophyseal processes of the caudal vertebrae with very elongated chevrons that are paired in double bony rods extending forward; ischium with large triangular obturator process situated above the ischial end; the pubis is propubic; stocky and elongated pubis with anteroposterior directed large distal expansion.[1]
According to the revised diagnosis by Turner et al. 2012, Achillobator can be distinguished based on the following combination of characteristics and autapomorphies: the promaxillary fenestra is completely exposed; the promaxillary and maxillary fenestra are elongate and vertically oriented at same level in the maxilla; metatarsal III is wide proximally; the femur is longer than the tibia; the pelvis is propubic; the obturator process on the ischium is large and triangular, and is situated on the proximal half of ischial shaft; the boot at distal symphysis of pubis is both cranially and caudally developed.[2]
Classification

Achillobator is classified as a dromaeosaurid, a group of very bird-like dinosaurs. In its original description, Achillobator was placed as a close relative of Dromaeosaurus.[1] It is part of the Eudromaeosauria, they were obligate carnivores, better known as the "true dromaeosaurids". Eudromaeosauria was first defined as a node-based clade by Nick Longrich and Philip J. Currie in 2009, as the most inclusive natural group containing Dromaeosaurus, Velociraptor, Deinonychus, and Saurornitholestes, their most recent common ancestor and all of its other descendants. The various "subfamilies" have also been re-defined as clades, usually defined as all species closer to the groups namesake than to Dromaeosaurus or any namesakes of other sub-clades.[16] The recent phylogenetic analysis, conducted by Hartman et al. 2019, surprisingly recovers Achillobator as the sister taxon of Yixianosaurus.[5]
During the description of Dakotaraptor in 2015, it was proposed a new cladistic analysis using data from the Theropod Working Group. Achillobator was found to be a relative of Utahraptor:[17]
| Eudromaeosauria |
| ||||||||||||||||||||||||||||||||||||||||||

Below are the results for the Eudromaeosauria phylogeny based on the phylogenetic analysis conducted by Currie and Evans in 2019. Achillobator was again recovered in the traditional Utahraptor+Achillobator clade, this time however, as a velociraptorine:[6]
| Eudromaeosauria |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paleobiology

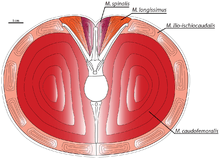
The structure of the hindlimbs and pelvic region of Achillobator indicates that the animal had massive thighs, with legs suited for moderate, fast-running. However, the pelvis of Achillobator has its own specialized femoral muscle retractors that lead to the suggestion of the unlikely locomotion method based on kangaroo-like jumps, or a similar way in which Archaeopteryx could have move.[1]
.jpg)
Manning et al. 2009 tested the function of the "sickle claw" of dromaeosaurids by analyzing the biomechanics of how stresses and strains would be distributed along the claws and into the limbs, and using comparisons within the curvature of the dromaeosaurid "sickle claw" on the foot with curvature in modern birds and mammals. They found that they were ideal for climbing and for a ground-dwelling life style.[18] Peter Mackovicky stated that the analysis might be correct on primitive dromaeosaurids (such as Microraptor) being tree-climbers, however, this doesn't explain why the giant Achillobator and Utahraptor (and other later dromaeosaurids) retained "sickle claws" when they were way too large to have climbed trees. Mackovicky suggested that larger dromaeosaurids adapted the claw to be used exclusively for a more aggressive predatory behaviour.[19]
The striking resemblance between the feet and legs of dromaeosaurids and those of accipitrid birds of prey, led Fowler et al. 2011 to propose that dromaeosaurids hunted in a similar way to those raptorial birds. They found that the feet and legs of dromaeosaurs resemble those of eagles and hawks by having an enlarged second claw and a similar range of flexion, but the metatarsals share more resemblance to those of owls. The model, RPR (Raptor Prey Restraint), proposes that dromaeosaurs leaped into their prey, immobilizing it with their body weight, and then hold it tightly with the large, sickle-shaped claws; afterwards, the dromaeosaur would start to feed on the animal while it's still alive and the death will eventually came from blood loss and organ failure. The arms or "wings", that were likely covered in long feathers (evidenced on Dakotaraptor[9], Velociraptor[7] and Zhenyuanlong[8]), may have been flapped by the dromaeosaur in order to stabilize its balance while restraining prey, along with this, the long, feathered tail probably worked as a counter-balance to the main body. Lastly, the snout would have been useful for finishing off its prey. With these observations, they established that Dromaeosaurids and Troodontids were niche partitioned as large and small prey predators, respectively.[20]
Paleoecology

The remains of Achillobator were unearthed from the Burkhant locality at the Bayan Shireh Formation.[1] This formation has been divided into Upper and Lower beds, with some localities representing lower and upper Cretaceous boundary.[21][22][23][24] Burkhant, has been represented as a Upper Cretaceous section, indicating that is part of the Upper Bayan Shireh.[25] Definitive age assignment for this formation is uncertain, with various hypotheses; based on comparative analyses, the Bayan Shireh fauna seems to correspond best with the Turonian through early Campanian stages of the Late Cretaceous, about 93 million to 80 million years ago.[26] However, the examination of the magnetostratigraphy of the formation seems to confirm that the entire Bayan Shireh lies within the Cretaceous Long Normal, which lasted only until the end of the Santonian stage, giving a Cenomanian-Santonian age, or, 98.5 million and 83.5 million years ago.[27] Nevertheless, according to Averianov and Sues in 2012, diverse biostratigraphic occurrences within formations in the Gobi Desert are indicatives of a correlation between the Upper Bayan Shireh and the Iren Dabasu Formation, approximately between 97 million and 83 million years ago.[24] Recent calcite U–Pb analyses have confirm the age of the Bayan Shireh Formation from 102 million to 86 million years ago.[28]
While other dromaeosaurids filled a variety of specialized ecological niches, mainly those of small predators or larger fish-eating forms, eudromaeosaurs, such as the large Achillobator, filled the niche of large-bodied predators of often medium to large-sized prey.[16] Therizinosaurs were the most abundant theropods in biodiversity across the formation, as evidenced on the remains of Enigmosaurus, Erlikosaurus and Segnosaurus.[29] Apparently, there was niche partitioning among species, such is the case of Erlikosaurus and Segnosaurus,[30] or Talarurus and Tsagantegia.[31]
Environment and paleoflora

The presence of caliche, fluvial and lacustrine sedimentation at various localities are indicators of a semi-arid climate with large water bodies, such as rivers, lakes and streams.[32][33][34][35] The Bayan Shireh formation was considered to be partially lacustrine by Samoilov and Benjamini in 1996[35], however, large scale cross-stratifications on various sandstone layers at the Baynshire locality are indicators of meandering fluvial systems; in addition to this, the presence of large river bodies drained the eastern part of the Gobi Desert.[27] Nevertheless, fluvial environments were again suggested by Averianov and Sues.[24] Cornaceae has been reported from the Khara Khutul locality, with Bothrocaryum gobience and Nyssoidea mongolica as representatives.[21] Additional findings on this locality and Bor Guvé seem to reflect the large presence of Angiosperm plants across the formation, with numerous fossil fruits recovered. These fruits are similar to Abelmoschus esculentus (Okra), however, the taxonomic affinities are quite unclear.[36]
Contemporaneous paleofauna
Paleofauna that were contemporaneous with the large dromaeosaurid in the Upper Bayan Shireh (or Late Cretaceous beds), included a rich diversity of dinosaur and non-dinosaur genera. Fellow dinosaur genera included the relatively large theropods Alectrosaurus[37], Erlikosaurus[38], Garudimimus[39] and Segnosaurus.[40] The heavy-built ankylosaurs Talarurus[41] and Tsagantegia.[42] Small marginocephalians Amtocephale[43] and Graciliceratops.[44] The basal hadrosauroid Gobihadros[25] and the large, long-necked titanosaur Erketu.[36]
Non-dinosaur taxa was present as well in the Upper Bayan Shireh, compromising semiaquatic and terrestrial reptiles, such as the turtles Lindholmemys[45], Gobiapalone and Trionyx[46], in addition, the crocodylomorph Paralligator.[47] Fishes are also known from the formation; the shark Hybodus and the remains of osteichthyan fish are fine representatives.[24] Several unnamed/indeterminate remains have been reported from localities across the formation, mostly dinosaur taxa: U. Azhdarchid(s)[48], U. Therizinosaur[49], U. Titanosaur[50] and an U. Velociraptorine-grade dromaeosaurid(s).[51][52][2][53]

See also
| Wikimedia Commons has media related to Achillobator. |
- Bayan Shireh Formation
- Eudromaeosauria
- Santonian
- List of Asian dinosaurs
- Timeline of dromaeosaurid research
References
- Perle, A.; Norell, M.A.; Clark, J. (1999). "A new maniraptoran Theropod−Achillobator giganticus (Dromaeosauridae)−from the Upper Cretaceous of Burkhant, Mongolia". Contributions from the Geology and Mineralogy Chair, National Museum of Mongolia (101): 1–105. OCLC 69865262.
- Turner, A.H.; Makovicky, P.J.; Norell, M.A. (2012). "A Review of Dromaeosaurid Systematics and Paravian Phylogeny". Bulletin of the American Museum of Natural History. 2012 (371): 1–206. doi:10.1206/748.1. hdl:2246/6352.
- Burnham, D.A.; Derstler, K.L.; Currie, P.J.; Bakker, R.T.; Zhou, Z.; Ostrom, J.H. (2000). "Remarkable New Birdlike Dinosaur (Theropoda: Maniraptora) from the Upper Cretaceous of Montana". The University of Kansas Paleontological Contributions. 13: 1–14. doi:10.17161/PCNS.1808.3761. ISSN 1046-8390.
- Norell, M.A.; Makovicky, P.J. (2004). "Dromaeosauridae". In Weishampel, D.B.; Dodson, P.; Osmólska, H. (eds.). The Dinosauria (2nd ed.). University of California Press. p. 207. ISBN 9780520941434.
- Hartman, S.; Mortimer, M.; Wahl, W.R.; Lomax, D.R.; Lippincott, J.; Lovelace, D.M. (2019). "A new paravian dinosaur from the Late Jurassic of North America supports a late acquisition of avian flight". PeerJ. 7: e7247. doi:10.7717/peerj.7247. PMC 6626525. PMID 31333906.
- Currie, P. J.; Evans, D. C. (2020). "Cranial Anatomy of New Specimens of Saurornitholestes langstoni (Dinosauria, Theropoda, Dromaeosauridae) from the Dinosaur Park Formation (Campanian) of Alberta". The Anatomical Record. 303 (4): 691–715. doi:10.1002/ar.24241. PMID 31497925.
- Turner, A.H.; Makovicky, P.J.; Norell, M.A. (2007). "Feather quill knobs in the dinosaur Velociraptor". Science. 317 (5845): 1721. Bibcode:2007Sci...317.1721T. doi:10.1126/science.1145076. PMID 17885130.
- Lü, J.; Brusatte, S.L. (2015). "A large, short-armed, winged dromaeosaurid (Dinosauria: Theropoda) from the Early Cretaceous of China and its implications for feather evolution". Scientific Reports. 11775 (5): 11775. doi:10.1038/srep11775. PMC 4504142. PMID 26181289.
- DePalma, R.A.; Burnham, D.A.; Martin, L.D.; Larson, P.L.; Bakker, R.T. (2015). "The First Giant Raptor (Theropoda: Dromaeosauridae) from the Hell Creek Formation". Paleontological Contributions. 2015 (14): 1–16. doi:10.17161/paleo.1808.18764.
- Turner, A.H.; Pol, D.; Clarke, J.A.; Erickson, G.M.; Norell, M.A. (2007). "A Basal Dromaeosaurid and Size Evolution Preceding Avian Flight". Science. 317 (5843): 1378–1381. doi:10.1126/science.1144066. PMID 17823350. Supporting Online Material
- Novas, F.E.; Pol, D.; Canale, J.I.; Porfiri, J.D.; Calvo, J.O. (2008). "A bizarre Cretaceous theropod dinosaur from Patagonia and the evolution of Gondwanan dromaeosaurids". Proceedings of the Royal Society B: Biological Sciences. 276 (1659): 1101–1107. doi:10.1098/rspb.2008.1554. ISSN 1471-2954. PMC 2679073. PMID 19129109.
- Holtz, T.R.; Rey, L.V. (2007). Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages. Random House. ISBN 9780375824197.CS1 maint: date and year (link) Genus List for Holtz 2012 Weight Information
- Paul, G.S. (2016). The Princeton Field Guide to Dinosaurs (2nd ed.). Princeton, New Jersey: Princeton University Press. p. 151. ISBN 9780691167664.
- Romer, A.S. (1923). "The pelvic musculature of saurischian dinosaurs" (PDF). Bulletin of the American Museum of Natural History. 48 (19): 605–617.
- Perle, A. (1998). "Primitive models of the pelvic and hindlimb structures-muscles of reptiles (lizards, crocodiles and birds)". Mongolian Geoscientist. 8 (10): e78574. doi:10.1371/journal.pone.0078574. PMC 3812989. PMID 24205268.
- Longrich, N.R.; Currie, P.J. (2009). "A microraptorine (Dinosauria–Dromaeosauridae) from the Late Cretaceous of North America". Proceedings of the National Academy of Sciences. 106 (13): 5002–5007. doi:10.1073/pnas.0811664106. PMC 2664043. PMID 19289829.
- DePalma, R.A.; Burnham, D.A.; Martin, L.D.; Larson, P.L.; Bakker, R.T. (2015). "The First Giant Raptor (Theropoda: Dromaeosauridae) from the Hell Creek Formation". Paleontological Contributions (14). doi:10.17161/paleo.1808.18764.
- Manning, P.L.; Margetts, L.; Johnson, M.R.; Withers, P.J.; Sellers, W.I.; Falkingham, P.L.; Mummery, P.M.; Barrett, P.M.; Raymont, D.R. (2009). "Biomechanics of dromaeosaurid dinosaur claws: Application of X-ray microtomography, nanoindentation, and finite element analysis". The Anatomical Record. 292 (9): 1397–1405. doi:10.1002/ar.20986. PMID 19711472.
- "Killing claws of Velociraptor… are for climbing". New Scientist. 203 (2725): 10. 2009. doi:10.1016/s0262-4079(09)62381-2.
- Fowler, D.W.; Freedman, E.A.; Scannella, J.B.; Kambic, R.E. (2011). "The Predatory Ecology of Deinonychus and the Origin of Flapping in Birds". PLOS ONE. 6 (12): e28964. Bibcode:2011PLoSO...628964F. doi:10.1371/journal.pone.0028964. PMC 3237572. PMID 22194962.
- Khand, Y.; Badamgarav, D.; Ariunchimeg, Y.; Barsbold, R. (2000). "Cretaceous system in Mongolia and its depositional environments". Cretaceous Environments of Asia. Developments in Palaeontology and Stratigraphy. 17. pp. 49–79. doi:10.1016/s0920-5446(00)80024-2. ISBN 9780444502766.
- Shuvalov, V.F. (2003). "The Cretaceous stratigraphy and palaeobiogeography of Mongolia". The Age of Dinosaurs in Russia and Mongolia. Cambrige University Press. pp. 256–278. ISBN 9780521545822.
- Eberth, D.A.; Kobayashi, Y.; Lee, Y.N.; Mateus, O.; Therrien, F.; Zelenitsky, D.K.; Norell, M.A. (2009). "Assignment of Yamaceratops dorngobiensis and associated redbeds at Shine Us Khudag (eastern Gobi, Dorngobi Province, Mongolia) to the redescribed Javkhlant Formation (Upper Cretaceous)". Journal of Vertebrate Paleontology. 29 (1): 295–302. doi:10.1080/02724634.2009.10010384.
- Averianov, A.; Sues, H. (2012). "Correlation of Late Cretaceous continental vertebrate assemblages in Middle and Central Asia" (PDF). Journal of Stratigraphy. 36 (2): 462–485.
- Tsogtbaatar, K.; Weishampel, D.B.; Evans, D.C.; Watabe, M. (2019). "A new hadrosauroid (Dinosauria: Ornithopoda) from the Late Cretaceous Baynshire Formation of the Gobi Desert (Mongolia)". PLOS ONE. 14 (4): e0208480. Bibcode:2019PLoSO..1408480T. doi:10.1371/journal.pone.0208480. PMC 6469754. PMID 30995236.
- Jerzykiewicz, T.; Russell, D.A. (1991). "Late Mesozoic stratigraphy and vertebrates of the Gobi Basin". Cretaceous Research. 12 (4): 345–377. doi:10.1016/0195-6671(91)90015-5. ISSN 0195-6671.
- Hicks, J.F.; Brinkman, D.L.; Nichols, D.J.; Watabe, M. (1999). "Paleomagnetic and palynologic analyses of Albian to Santonian strata at Bayn Shireh, Burkhant, and Khuren Dukh, eastern Gobi Desert, Mongolia". Cretaceous Research. 20 (6): 829–850. doi:10.1006/cres.1999.0188.
- Kurumada, Y.; Aoki, S.; Aoki, K.; Kato, D.; Saneyoshi, M.; Tsogtbaatar, K.; Windley, B. F.; Ishigaki, S. (2020). "Calcite U–Pb age of the Cretaceous vertebrate‐bearing Bayn Shire Formation in the Eastern Gobi Desert of Mongolia: usefulness of caliche for age determination". Terra Nova. doi:10.1111/ter.12456.
- Lee, Y.M.; Lee, H.J.; Kobayashi, Y.; Carabajal, A.P.; Barsbold, R.; Fiorillo, A.R.; Tsogtbaatar, K. (2019). "Unusual locomotion behaviour preserved within a crocodyliform trackway from the Upper Cretaceous Bayanshiree Formation of Mongolia and its palaeobiological implications". Palaeogeography, Palaeoclimatology, Palaeoecology. 533 (109353): 2. doi:10.1016/j.palaeo.2019.109239.
- Zanno, L.E.; Tsogtbaatar, K.; Chinzorig, T.; Gates, T.A. (2016). "Specializations of the mandibular anatomy and dentition of Segnosaurus galbinensis (Theropoda: Therizinosauria)". PeerJ. 4: e1885. doi:10.7717/peerj.1885. PMC 4824891. PMID 27069815.
- Park, J.Y.; Lee, Y.N.; Currie, P.J.; Kobayashi, Y.; Koppelhus, E.; Barsbold, R.; Mateus, O.; Lee, S.; Kim, S.H. (2020). "Additional skulls of Talarurus plicatospineus (Dinosauria: Ankylosauridae) and implications for paleobiogeography and paleoecology of armored dinosaurs". Cretaceous Research. 108: 104340. doi:10.1016/j.cretres.2019.104340.
- Vasiliev, V.G.; Volkhonin, V.C.; Grishin, G.L.; Ivanov, A.K.; Marinov, I.A.; Mokshancev, K.B. (1959). Geological structure of the People's Republic of Mongolia (stratigraphy and tectonics). p. 492.
- Sochava, A.V. (1975). "Stratigraphy and lithology of the Upper Cretaceous sediments in southern Mongolia. In Stratigraphy of Mesozoic sediments of Mongolia". Transactions of Joint Soviet–Mongolian Scientific Research and Geological Expedition. 13: 113–182.
- Martinson, G.G. (1982). "The Upper Cretaceous mollusks of Mongolia". Sovmestnaya Sovetsko– Mongolskaya Paleontolog-icheskaya Ekspeditsia. 17: 5–76.
- Samoilov, V.S.; Benjamini, C. (1996). "Geochemical features of dinosaur remains from the Gobi Desert, South Mongolia". PALAIOS. 11 (6): 519–531. doi:10.2307/3515188. JSTOR 3515188.
- Ksepka, D.T.; Norell, M.A. (2006). "Erketu ellisoni, a long-necked sauropod from Bor Guvé (Dornogov Aimag, Mongolia)" (PDF). American Museum Novitates. 2006 (3508): 1–16. doi:10.1206/0003-0082(2006)3508[1:EEALSF]2.0.CO;2.
- Perle, A. (1977). "O pervoy nakhodke Alektrozavra (Tyrannosauridae, Theropoda) iz pozdnego Mela Mongolii" [On the first discovery of Alectrosaurus (Tyrannosauridae, Theropoda) in the Late Cretaceous of Mongolia]. Shinzhlekh Ukhaany Akademi Geologiin Khureelen (in Russian). 3 (3): 104–113.
- Barsbold, R.; Perle, A. (1980). "Segnosauria, a new suborder of carnivorous dinosaurs" (PDF). Acta Palaeontologica Polonica. 25 (2): 190–192.
- Barsbold, R. (1981). "Toothless carnivorous dinosaurs of Mongolia" (PDF). Transactions, Joint Soviet–Mongolian Palaeontological Expedition. 15: 28–39.
- Perle, A. (1979). "Segnosauridae — novoe semejstvo teropod iz pozdnego mela Mongolii" [Segnosauridae — a new family of theropods from the Late Cretaceous of Mongolia] (PDF). Transactions of the Joint Soviet-Mongolian Paleontological Expedition (in Russian). 8: 45–55.
- Maleev, E.A. (1952). "Noviy ankilosavr is verchnego mela Mongolii" [A new ankylosaur from the Upper Cretaceous of Mongolia] (PDF). Doklady Akademii Nauk SSSR (in Russian). 87 (2): 273–276.
- Tumanova, T.A. (1993). "O novom pantsirnov dinozavre iz iugo-vostochnoy Gobi" [A new armored dinosaur from Southeastern Gobi]. Paleontologicheskii Zhurnal (in Russian). 27 (2): 92–98.
- Watabe, M.; Tsogtbaatar, K.; Sullivan, R.M. (2011). "A new pachycephalosaurid from the Baynshire Formation (Cenomanian-late Santonian), Gobi Desert, Mongolia" (PDF). Fossil Record 3. New Mexico Museum of Natural History and Science, Bulletin. 53: 489–497.
- Sereno, P.C. (2000). "The fossil record, systematics and evolution of pachycephalosaurs and ceratopsians from Asia" (PDF). The Age of Dinosaurs in Russia and Mongolia. Cambridge University Press. pp. 489–491.
- Sukhanov, V.B.; Danilov, I.G.; Syromyatnikova, E.V. (2008). "The Description and Phylogenetic Position of a New Nanhsiungchelyid Turtle from the Late Cretaceous of Mongolia". Acta Palaeontologica Polonica. 53 (4): 601–614. doi:10.4202/app.2008.0405.
- Danilov, I.G.; Hirayama, R.; Sukhanov, V.B.; Suzuki, S.; Watabe, M.; Vitek, N.S. (2014). "Cretaceous soft-shelled turtles (Trionychidae) of Mongolia: new diversity, records and a revision". Journal of Systematic Palaeontology. 12 (7): 799–832. doi:10.1080/14772019.2013.847870.
- Turner, A.H. (2015). "A Review of Shamosuchus and Paralligator (Crocodyliformes, Neosuchia) from the Cretaceous of Asia". PLOS ONE. 10 (2): e0118116. doi:10.1371/journal.pone.0118116. PMC 4340866. PMID 25714338.
- Watabe, M.; Suzuki, D.; Tsogtbaatar, K. (2009). "The first discovery of pterosaurs from the Upper Cretaceous of Mongolia" (PDF). Acta Palaeontologica Polonica. 54 (2): 231–242. doi:10.4202/app.2006.0068.
- Kobayashi, Y.; Chinzorig, T.; Tsogtbaatar, K.; Barsbold, R. (2015). "A new therizinosaur with functionally didactyl hands from the Bayanshiree Formation (Cenomanian-Turonian), Omnogovi Province, southeastern Mongolia" (PDF). Journal of Vertebrate Paleontology. Program and Abstracts.
- Ksepka, D.; Norell, M.A. (2010). "The Illusory Evidence for Asian Brachiosauridae: New Material of Erketu ellisoni and a Phylogenetic Reappraisal of Basal Titanosauriformes" (PDF). American Museum Novitates. 3700 (3700): 1–27. doi:10.1206/3700.2.
- Currie, P.J.; Varricchio, D.J. (2004). "A new dromaeosaurid from the Horseshoe Canyon Formation (Upper Cretaceous) of Alberta, Canada". Feathered Dragons. Indiana University Press. p. 128. ISBN 0253343739. Book preview
- Kubota, K.; Barsbold, R. (2007). "New dromaeosaurid (Dinosauria Theropoda) from the Upper Cretaceous Bayanshiree Formation of Mongolia". Journal of Vertebrate Paleontology. 27 (suppl. to 3): 102A. doi:10.1080/02724634.2007.10010458.
- Kubota, K. (2015). "Descriptions of Mongolian Dromaeosaurids (Dinosauria: Theropoda) and phylogeny of Dromaeosauridae". PHD Dissertation (in Japanese).
Paraves | ||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||












{kind=link}