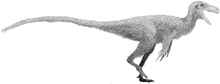
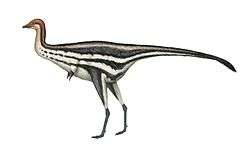
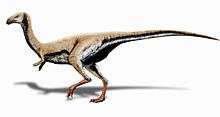
Limusaurus
Limusaurus is a genus of theropod dinosaur that lived in what is now western China during the Late Jurassic, around 161 to 157 million years ago. The genus and species Limusaurus inextricabilis was described in 2009 from specimens found in the Upper Shishugou Formation in the Junggar Basin. The genus name consists of the Latin words for "mud" and "lizard", and the species name means "impossible to extricate", both referring to these specimens possibly dying after being mired. Limusaurus was a small, slender animal, about 1.7 metres (5 ft 7 in) in length, that had a long neck and legs but also very small forelimbs (with reduced first and fourth fingers). It underwent a drastic morphological transformation as it aged; while juveniles were toothed, these teeth were completely lost and replaced by a beak with age, features convergently similar to the later ornithomimid theropods as well as the earlier non-dinosaurian shuvosaurids
| Limusaurus | |
|---|---|
 | |
| Skeletal diagram showing the preserved remains of the holotype specimen | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Clade: | Dinosauria |
| Clade: | Saurischia |
| Clade: | Theropoda |
| Family: | †Noasauridae |
| Subfamily: | †Elaphrosaurinae |
| Genus: | †Limusaurus Xu et al., 2009 |
| Species: | †L. inextricabilis |
| Binomial name | |
| †Limusaurus inextricabilis Xu et al., 2009 | |
Limusaurus is the first known member of the group Ceratosauria from Asia. Along with its closest relative, Elaphrosaurus, it was a member of the family Noasauridae, a group of small and lightly built abelisaurs. The pattern of digit reduction in Limusaurus has been used to support the contested hypothesis that the three-fingered hand of tetanuran theropods is the result of the loss of the first and fifth digits from the ancestral five-fingered theropod hand, which has implications for the evolution of birds, though this has been contested. The change to toothlessness in adult probably correspond to a dietary shift from omnivory to herbivory, which is also confirmed by the presence of gastroliths (stomach stones) in adults. SInce many specimens were found together, it is possible Limusaurus lived in groups. Its fossils were discovered in rocks dated to the Oxfordian age. Specimens of Limusaurus (along with other small animals) appear to have been mired in mud pits created by the footsteps of sauropod dinosaurs.
Discovery and naming

Between 2001 and 2006, a Chinese-American team of paleontologists examining the Wucaiwan locality in the Shishugou Formation, in the northeastern Junggar Basin of Xinjiang, China, discovered three bonebeds (numbered TBB 2001, TBB 2002, and TBB 2005, found by T. Yu and J. Mo). The bonebeds were dominated by the remains of small theropod dinosaurs, representing at least three genera, with most belonging to a small ceratosaur, the first member of this group found in Asia. Stacked skeletons from these bonebeds were removed from the field in blocks jacketed by plaster and encased in crates, and a cast of one block was also made, making it available for study after the specimens had been extracted from the original.[1][2]
In 2009, the small ceratosaur was described by paleontologist Xu Xing and colleagues, who named it Limusaurus inextricabilis. The genus name consists of the words limus, Latin for "mud" or "mire", and saurus, greek for "lizard", and the species name means "impossible to extricate"; both names refer to how these specimens appear to have died after being mired.[2] The name has also been translated as "mire lizard who could not escape".[3] The description incorporated data from two subadult specimens which are both stored at the Institute of Vertebrate Paleontology and Paleoanthropology (IVPP) in Beijing: the holotype (a subadult cataloged under the specimen number IVPP V 15923) is an almost complete and articulated (still connected) skeleton, missing only the hindmost tail vertebrae, and is preserved next to another specimen (a large juvenile, IVPP V 20098) which is missing the front part of the skeleton. The other (an adult, IVPP V 15304, inaccurately referred to by the number IVPP V 16134 in the original description) is a likewise almost complete and articulated specimen that is missing only the skull, and is 15% larger than the holotype.[2][4]
Seventeen additional Limusaurus specimens were described in 2016, excavated from the same blocks as those described in 2009. These specimens include six juveniles (one year in age or less), ten subadults (two-six years in age), and one adult (more than six years in age). Like the holotype and second specimen, these specimens are also stored at the Institute of Vertebrate Paleontology and Paleoanthropology.[4][1] The toothless adults and toothed juveniles were initially thought to be different kinds of dinosaurs, and were studied separately, until it was realized they represent the same species.[5]
Description

Limusaurus was a small and slender animal. The holotype (which was originally considered an adult based on the level of fusion of its bones, but later as a subadult when analyzed along with other specimens) is estimated to have been about 1.7 metres (5 ft 7 in) in length[2] and the weight of the animal has been estimated at about 15 kilograms (33 lb). One adult specimen is estimated to have been 15% larger than the holotype.[6][4] Its neck and legs were proportionally long; the latter were 1.8 times the length of its torso. Several features of the animal, such as the small head with large orbits, toothless jaws, and its long neck and legs, were very similar to those of the Cretaceous ornithomimid theropods, as well as the Triassic non-dinosaurian shuvosaurids, representing a significant case of convergent evolution among these three distinct groups of archosaurs.[2] While Limusaurus has sometimes been depicted with feathers and may have had them, there is no evidence of such structures.[7][3]
Skull
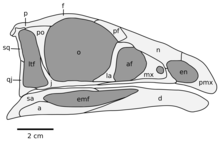
The skull was relatively tall and short, roughly half the length of the femur (upper thigh bone). The tip of its jaws was covered by a beak, a feature that was previously unknown in theropods more basal (or "primitive") than coelurosaurs, the group that includes the most bird-like dinosaurs. Uniquely to Limusaurus, the inner edge of the premaxilla, the frontmost bone of the upper jaw, was convex. The nasal bone was distinct in having a "shelf" on its side, was short, wide, less than one third of the length of the skull roof, and twice as long as it was wide. As in most dinosaurs, the skull featured five principal fenestrae (openings), including the bony nostril, eye opening, antorbital fenestra (between the nostril and eye), as well as the upper and lower temporal fenestra (on the top and on the side of the skull's rear, respectively). As in other ceratosaurians, parts of the bony nostril were formed by the maxilla (upper jaw bone); also, the antorbital fenestra was proportionally small, and the rear part of the nasal bone formed parts of the cavity which contained this opening. The external naris was large and located in a hindwards position, similar to tetanuran theropods. The eye opening was large, while the lateral temporal fenestra was not as large as would be expected from more derived members of the Ceratosauria. The lower part of the lacrimal, the bone that formed the front margin of the eye opening, was unique in being strongly inclined forwards. The jugal bone, which formed the floor of the eye opening, was slender, and its rami (or branches) were rod like, uniquely for this genus.[2][4]
Another opening, the mandibular fenestra of the lower jaw, was generally large in ceratosaurians. In Limusaurus, it was especially large, accounting for 40% of the length of the entire lower jaw, a distinguishing feature of the genus. The dentary (tooth-bearing bone at the front of the lower jaw) was short compared to the rest of the lower jaw, as in other ceratosaurians. The front end of the dentary was down-turned and had a convex inner margin, similar to the related Masiakasaurus. The angular bone of the lower jaw was positioned significantly forwards in relation to the hind end of the mandible, similar to other ceratosaurians. Juveniles had nine teeth in each side of the upper jaw, and twelve in each side of the lower; they were gradually lost as they grew, disappearing by adulthood.[2][4]
Postcranial skeleton
The cervicals (neck vertebrae) were elongated as in the closely related Elaphrosaurus. The axis (second neck vertebra) lacked the pleurocoel (excavation) on its front end and the foramina (openings) piercing its neural arch that are seen in derived ceratosaurians. As in other noasaurids, the neural spines of the cervicals were positioned more towards the front end of their vertebrae than is the case in other theropods.[2]
Distinctively, the scapula (shoulder blade) bore a prominent ridge at its front edge. It also had a comparatively high acromion process. The sternum was fused into a single, large, continuous plate, another feature that evolved independently in coelurosaurs (convergent evolution). Limusaurus also had a furcula, or wishbone, which previously was unknown among ceratosaurians. The head of the humerus (upper arm bone) was bulging, and the deltopectoral crest, a forward-directed bony flange of the humerus that served for muscle attachment, was long and angled; these features were typical for ceratosaurians. In the forearm, the radius was longer than the ulna, and the olecranon process, a bony extension on the upper end of the ulna that served for muscle attachment, was absent in Limusaurus. Both features are considered autapomorphies of the genus. As in other ceratosaurians, ossified wrist bones were absent.[2]
As is typical for abelisaurids, the arms and hands of Limusaurus were considerably reduced, even more so than in Ceratosaurus. Uniquely in Limusaurus, the first finger was missing entirely, and the first metacarpal was shorter than the other metacarpals. The second metacarpal was more robust than the other metacarpals, which is another autapomorphy of the genus. The second finger had three phalanges (finger bones). The third finger only had three phalanges as opposed to four in other early theropods. Although the fourth finger is not preserved, the tip of the fourth metacarpal indicates the presence of a joint and therefore the presence of a phalanx; it is likely that this was the only phalanx of the fourth finger.[2][8] The unguals (claw bones) were short, stout, and expanded at their base.[9] They had two grooves on their sides, a feature also found in Masiakasaurus.[2]
Of the pelvis, the ilium was small and tilted towards the midline of the body, as is the case in Elaphrosaurus. As in other ceratosaurians, the lower end ("boot") of the pubis was large and expanded. Unique for the genus, it pointed backwards in a hook-like shape and had a ridge on each side. The legs of Limusaurus had proportions that were well-adapted to running, with their lower segments much longer than the femur: the tibiotarsus, the fusion of the tibia (shin bone) and tarsal bones, was 1.2 times the length of the femur, and the foot was 1.3 times the length of the femur. The upper half of the femur was triangular in cross section, a feature shared with Masiakasaurus. The metatarsals of the other three toes were arranged in an arc, with the fourth metatarsal straight and adhering tightly to the third for its entire length; these features are unique to Limusaurus. Another unique feature, the hallux (the first toe) was reduced, being only 17% the length of the third metatarsal. As in other ceratosaurians, the unguals of the foot had two grooves on their sides.[2]
Classification
Limusaurus is a member of the Ceratosauria. It had several skull features in common with basal theropods such as other ceratosaurs and coelophysoids, but it also shared a number of traits, including the beak and the fused sternum, convergently with the later coelurosaurs. Originally, it was described as a basal ceratosaurian,[2] with both it and Elaphrosaurus being recovered as the most basal ceratosaurians in various phylogenetic analyses.[10][11][12] However, subsequent research has placed it in the more derived (or "advanced") group Abelisauroidea,[13] specifically as a member of the Noasauridae.[14] A 2017 analysis by paleontologist Shuo Wang and colelagues conducted in the study of multiple individuals of different ages found Limusaurus and Elaphrosaurus to group in the clade Elaphrosaurinae, within the family Noasauridae. Noasauridae was placed in a position outside Neoceratosauria, the group containing Ceratosaurus and Abelisauridae. All the juvenile Limusaurus specimens used in the study grouped together to the exclusion of adult specimens, showing that their anatomy changed significantly through growth.[4]
The cladogram below shows the position of Limusaurus within Noasauridae according to Wang and colleagues, 2017.[4]

| Noasauridae |
| ||||||||||||||||||||||||||||||||||||
A contrasting analysis conducted by Rauhut and Carrano in 2016 found slightly different relationships within Noasauridae. The specimen CCG 20011 (an as-of-yet unnamed taxon formerly referred to the tetanuran Chuandongocoelurus), which was not included in other analyses, was found to be closer to Elaphrosaurus than Limusaurus, with the three taxa representing Elaphrosaurinae. Velocisaurus, Noasaurus and Masiakasaurus were the only taxa within Noasaurinae, as Laevisuchus and Deltadromeus were placed basal to the group of Noasaurinae and Elaphrosaurinae within Noasauridae. The only specimen of Elaphrosaurus itself is missing its skull and hands among other elements, and its affinities were long unclear (it was often considered an ornithomimosaur well into the 1990s) until the more complete Limusaurus was found, which Rauhut and Carrano used to extrapolate the complete length of the former taxon at 6.0–6.3 m (19.7–20.7 ft).[14][15] A 2019 study by Max Langer and colleagues, which was based on the same data set used by the 2016 study, recovered the same interrelationships within Elaphrosaurinae.[16]
Limusaurus is the first definite ceratosaur from eastern Asia to be discovered.[17] It is also one of the earliest members of the group Ceratosauria, living during the Oxfordian stage of the Jurassic period (approximately 161-157 million years ago). Its discovery shows that the Asian dinosaurian fauna was less endemic during the Middle/Late Jurassic period than previously thought and suggests a possible land connection between Asia and other continents during that period.[2]
Digit homology
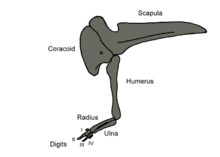
The most basal theropods have five digits on the hand. Along the lineage that led to birds, however, the number of digits in the hand decreased; by the emergence of the group Tetanurae, which includes birds, two digits had disappeared from the hand, leaving three digits. Traditionally, it has been hypothesized that the digits lost were the two outermost digits, i.e. digits IV and V, in a process known as Lateral Digit Reduction (LDR).[2][18]
However, the hypothesis of LDR is in contradiction to some embryological studies on birds which show that, from five developmental sites, the digits that develop are the three middle digits (II, III, IV). This inconsistency has been a matter of debate for almost 200 years,[19] and has been used by Alan Feduccia to support the controversial hypothesis that birds are descended not from theropods but from some other group of archosaurs which had lost the first and fifth digits.[20] To explain the discrepancy between morphological and embryological data in the context of bird origins, an alternative scenario to LDR was developed by Tony Thulborn and Tim Hamley in 1982. In this scenario, the digits I and V of theropods were reduced in the process of Bilateral Digit Reduction (BDR), with the remaining digits developing to resemble the former digits I-III.[2][21] Limusaurus was initially considered as evidence for the BDR hypothesis due to it—and other ceratosaurians—having a reduced first digit, with researchers hypothesizing that a similar pattern of reduction occurred among the tetanurans (the sister group of the ceratosaurians).[2][22]
Several additional hypotheses have been proposed to improve upon and reconcile the LDR and BDR hypotheses. One predominantly favored hypothesis, first developed by Günter P. Wagner and Jacques Gauthier in 1999,[23] involves a "frameshift" of the digits; the first digit fails to grow in the first developmental site due to not receiving the necessary signals, which has the effect of shifting digits I-III to the positions of II-IV. Thus, while digits I-III from the ancestral theropod are retained, they do not grow in the same location.[8][18][24][25][26][27] A version of the frameshift hypothesis modified to incorporate both elements of BDR and fossil evidence from Limusaurus and other theropods, the "thumbs down" hypothesis of Daniel Čapek and colleagues, suggests that this frameshift took place after the reduction of both the first and the fourth digits in the theropod lineage.[28] The major alternative hypothesis supported by Xing and colleagues, known as the "lateral shift hypothesis", considers a partial, step-wise frameshift in which, from a four-fingered hand with reduced digits I and IV, I fully disappears while IV develops into a fully-fledged finger, with II-IV taking on the morphologies of the former I-III.[2][18][29]
It is plausible that ceratosaurians underwent BDR independent of the tetanurans,[2] and therefore have no bearing on the issue of avian digit homology.[27] In particular, as demonstrated by teratological analysis, the abbreviation (mesomelia) and loss of digits (hypophalangia) in the forelimb of Limusaurus is likely the result of a developmental anomaly that appeared exclusively in and persisted throughout the evolutionary lineage of ceratosaurians, and is unconnected to the pattern of digital reduction and frameshift that occurred in tetanurans.[8] This is supported by the hands of the ceratosaurians Ceratosaurus, Berberosaurus, and Eoabelisaurus having plesiomorphic (i.e. more similar to the ancestral condition of theropods than to derived abelisaurs) metacarpal bones comparable to those of the more basal theropod Dilophosaurus.[30] The divergent developmental pathways of ceratosaurians and tetanurans is associated with a difference in forelimb function; tetanurans utilized their hands for grasping, while the hands of ceratosaurians almost certainly played no role in predation.[18]
Paleobiology
Growth

Analysis of growth rings (visible in bone cross-sections and analogous to the growth rings of trees) of the tibiae from the various developmental stages of Limusaurus indicates that the animal was skeletally mature at six years in age. In older specimens, the outermost growth rings are very close together (forming what is known as the external fundamental system), indicating that rapid growth had ceased in these particular individuals.[4]
Specimens of Limusaurus show 78 different anatomical changes that occurred as the animals grew. In particular, their heads became proportionally shallower, their middle hand bones lengthened, and the "hook" of their pubis grew longer. The shaft of the quadrate bone in the skull also straightened in adults, and the tips of their lower jaws became more downturned.[4]
The most obvious change that happened during the growth of Limusaurus is the complete loss of teeth from juveniles to adults. Juveniles began with one tooth in the premaxilla, eight in the maxilla, and eleven in the lower jaw (42 teeth in total). At the next stage, the first, sixth, and eighth teeth in the maxilla, as well as the sixth in the lower jaw have all been lost (34 teeth in total), although the sockets are still present, and there is a small replacement tooth in the socket of the sixth lower tooth. During this stage, use of teeth and normal tooth replacement likely ceased or became reduced, since none of the still-erupted teeth bear any wear marks or resorption. As the specimens grew, the transformation became more radical. In subadult and adult specimens older than one year in age, all the teeth are missing. CT scanning shows that only five empty tooth sockets remain in the adult maxilla; all the sockets in the lower jaw have fused into a single, hollow canal, and the rest of the tooth sockets have been obliterated.[4]
The loss of teeth with age in Limusaurus is the most extreme case of tooth morphology changing with age among dinosaurs. In fact, Limusaurus is one of the only known jawed vertebrates where teeth are completely lost during growth (the others are the red mullet and striped red mullet, several armored catfish, and the platypus). Its complicated pattern of losing teeth from both the front and the back is most similar to that of the avialan Jeholornis.[31][32] The early halt in tooth replacement possibly resulted from the regression of the replacement tooth buds during the first year, as in the veiled chameleon.[33][32] The replacement of teeth by a beak during growth in Limusaurus suggests that beaks in other lineages of theropods, and indeed beaked animals in general, may have evolved heterochronically, i.e. that beaks first occurred in adults and then gradually extended to juveniles. This is also indicated by the retention of the developmental signal pathways necessary to induce tooth development in birds.[34][4][32]
Feeding and diet

Adult specimens of Limusaurus show morphological traits associated with herbivory that are shared with other groups of herbivorous theropods, including the Ornithomimosauria, Therizinosauria, Oviraptorosauria, and Alvarezsauroidea, along with the troodontid Jinfengopteryx.[35] In particular, adult specimens are characterized by the presence of gastroliths (stomach stones), with older adults having gastroliths that are larger and more numerous than those of younger adults. The size and quantity of these gastroliths are comparable to those of birds in young adults, and those of ornithomimosaurs and oviraptorosaurs in old adults. These groups of theropods all used gastroliths in processing plant matter, suggesting that Limusaurus did the same; the increased amount of gastroliths in older adults may be indicative of the gut's ability to process plant matter more finely as they aged. An isotope analysis of the available specimens of Limusaurus specimens likewise showed that adults consistently match the isotope signatures of other herbivorous dinosaurs. In the same analysis, juveniles and subadults were found to vary greatly in their isotopic signatures; this indicates that juveniles were likely omnivorous (feeding on both animals and plants), but switched to strict herbivory as they aged.[4] This is comparable to the diet shift experienced by the aforementioned mullets and armored catfish.[4]
Social behavior
The paleontologist David A. Eberth and colleagues suggested in 2010 that the large number of Limusaurus specimens in the Shishugou Formation mud pits suggests they were either abundant among the small vertebrate animals in the area, or that the trapped individuals had been drawn there. They found it unlikely that animals were trapped on any other basis than size, and pointed out that it was difficult to explain why herbivores like Limusaurus would be attracted to sites where other animals were mired, so they considered it most likely that the larger number of Lumusaurus was accidental; the time of miring would have taken weeks to months. These authors also suggested that the abundance of Lumusaurus inferred for the area and the evidence for successive, rapid burials of each individual made it possible that Lumusaurus was gegarious, living in goups. There is some evidence for gregariousness in many small theropods and that there may have been social behaviours related to age, but it is yet unknown whether the bone beds cotaining Limusaurus specimens preserve evidence of sociality and segregation related to age.[1] Paleontologist Rafael Delcourt agreed in 2018 that assemblages of associated Limusaurus and Masiakasaurus specimens suggests these small ceratosaurs lived in groups.[36]
Paleoenvironment

All known fossils were recovered from the Shishugou Formation, a succession of sedimentary rocks that was deposited at the northeastern margin of the Junggar foreland basin and is ca. 350–400 m in thickness. Limusaurus occurs in the upper part of the formation, which represents a variety of environments, including alluvial fans and alluvial plains, streams, wetlands, and shallow lakes. During the time when Limusaurus lived, the environment would have been consistently warm and dry, judging by the abundance of coal and carbon-rich deposits.[1][37] The climate was probably highly seasonal due to monsoonal influences, with warm, wet summers and dry winters.[38] The climate enabled the growth of a richly forested environment; the forest would have been dominated by Araucaria trees, with the undergrowth being occupied by Coniopteris tree ferns, Anglopteris and Osmunda ferns, Equisetites horsetails, and Elatocladus shrubs.[39]
The environment of the Shishugou Formation hosted a diverse assemblage of animals. More than 35 species of vertebrates are known from fossils, including at least 14 dinosaur species.[1] Contemporaries of Limusaurus in the Wucaiwan locality include the theropods Haplocheirus, Zuolong, and Guanlong, the latter of which is, like Limusaurus, frequently found in mud deposits;[1] the sauropod Mamenchisaurus; the ornithischians Gongbusaurus and Yinlong; the tritylodontid Yuanotherium; the mammal Acuodulodon; the crocodyliforms Sunosuchus, Junggarsuchus, and Nominosuchus; and the turtles Xinjiangchelys and Annemys.[40]
Taphonomy

The known fossil material of Limusaurus consists of large assemblages of individuals mired in mud pits. Two of the mud pits containing Limusaurus specimens were found at the same stratigraphic level, while a third was some 6.5 metres (21 ft) higher in the stratigraphic column.[4] The mud pits are around 2 m wide and range in depth from 1–2 metres (3 ft 3 in–6 ft 7 in). The mud pits occur within distinctive rock layers that can be followed for hundreds of meters to kilometers. These layers, though not the pits themselves, comprise pale-colored laminated successions of mudstone 0.5 to 2 m in thickness. The mudstones probably accumulated either in standing water or a water-saturated substrate; root traces at the top of the layers indicate a marsh-like environment. One of the layers is topped by a crust-like mudstone layer that indicates soil formation and exposure to air. Volcanic minerals found within the mudstone layers indicate volcanic events during deposition. The mud pits themselves show a tan-colored, silty to sandy claystone with abundant plant fragments and root traces. At their margins, the laminae of the surrounding mudstone are deformed or truncated, suggesting that the mudstone was partly fluid, plastic, and brittle at the time when the skeletons were deposited. Fragments of the crust topping one of the mudstone layers were found within the pits, indicating that the crust collapsed downwards into the pits.[1]
The deformation structures and the consistent size of the pits suggest that they represent the footprints of giant sauropods such as Mamenchisaurus, which was likewise found in the Shishugou Formation and would have had a mass of over 20 metric tons (22 short tons) and a limb length of over 3 metres (9.8 ft). Other possible explanations, including sand volcanoes or sinkholes, can be ruled out because characteristic sedimentological features are lacking. The possibility that the encased dinosaurs could have created the pits themselves can likewise be ruled out given their small size; the largest dinosaur found in the pits, Guanlong, would merely have been 66 centimetres (26 in) tall. When creating the pits, the footsteps of the giant dinosaurs could have lead to soil liquefaction, creating a trap for smaller animals including Limusaurus. The size of these assemblages can be attributed to the tendency of smaller animals to become trapped in the mud.[1]
Limusaurus is the most abundant dinosaur found in the mud pits. One of the three pits, TBB2001, contained four Limusaurus individuals while other species are absent. TBB2002, on the other hand, contained four theropod dinosaur skeletons including two Limusaurus, two Guanlong and one individual of a not yet described species. The third pit, TBB2005, contained at least nine Limusaurus individuals but also the tail of a small ornithischian dinosaur as well as two crocodyliforms, two mammals, a turtle and three tritylodontid cynodonts. The completeness of the skeletons is variable; at least half of the theropod skeletons have been complete skeletons, with missing parts due to recent erosion. Rear parts of the skeletons tend to be more common than front parts. Most individuals were embedded laying on their sides, though some lie on their backs or undersides. In pits TBB2001 and TBB2002 the skeletons lie one above the other. Bones pertaining to the same individual are often lying upon each other in direct contact, while there is no direct contact between bones of separate individuals. These observations lead Eberth and colleagues to conclude that the skeletons must have accumulated within the mud pits over an extended time span rather than during a short-term death event. The pits were referred to as "dinosaur death pits" in the scientific paper that described them and in media reporting.[1][41][5]
References
- Eberth, D.A.; Xing, X.; Clark, J.A. (2010). "Dinosaur death pits from the Jurassic of China". PALAIOS. 25 (2): 112–125. Bibcode:2010Palai..25..112E. doi:10.2110/palo.2009.p09-028r. Archived from the original on December 30, 2016. Retrieved December 30, 2016.
- Xu, X.; Clark, J.M.; Mo, J.; Choiniere, J.; Forster, C.A.; Erickson, G.M.; Hone, D.W.E.; Sullivan, C.; Eberth, D.A.; Nesbitt, S.; Zhao, Q.; Hernandez, R.; Jia, C.-K.; Han, F.-L.; Guo, Y. (2009). "A Jurassic ceratosaur from China helps clarify avian digital homologies" (PDF). Nature. 459 (18): 940–944. Bibcode:2009Natur.459..940X. doi:10.1038/nature08124. PMID 19536256. Archived (PDF) from the original on September 22, 2017. Retrieved July 13, 2019.
- "Beaked, Bird-like Dinosaur Tells Story of Finger Evolution". www.nsf.gov. National Science Foundation. Retrieved May 28, 2020.
- Wang, S.; Stiegler, J.; Amiot, R.; Wang, X.; Du, G.-H.; Clark, J.M.; Xu, X. (2017). "Extreme Ontogenetic Changes in a Ceratosaurian Theropod" (PDF). Current Biology. 27 (1): 144–148. doi:10.1016/j.cub.2016.10.043. PMID 28017609.
- Hunt, K. (2016). "Why do birds have beaks and not teeth? This dino may have the answer". CNN. Retrieved June 26, 2020.
- Paul, G.S. (2016). "Theropods". The Princeton Field Guide to Dinosaurs (2nd ed.). Princeton: Princeton University Press. p. 82.
- "Pecking order: toothless dinosaur points way to evolution of the beak". The Guardian. December 23, 2016. Retrieved May 28, 2020.
- Guinard, G. (2016). "Limusaurus inextricabilis (Theropoda: Ceratosauria) gives a hand to evolutionary teratology: a complementary view on avian manual digits identities". Zoological Journal of the Linnean Society. 176 (3): 674–685. doi:10.1111/zoj.12329.
- Agnolin, F.L.; Chiarelli, P. (2010). "The position of the claws in Noasauridae (Dinosauria: Abelisauroidea) and its implications for abelisauroid manus evolution". Paläontologische Zeitschrift. 84 (2): 293–300. doi:10.1007/s12542-009-0044-2.
- Carrano, M.T.; Sampson, S.D. (2008). "The Phylogeny of Ceratosauria (Dinosauria: Theropoda)". Journal of Systematic Palaeontology. 6 (2): 183–236. doi:10.1017/S1477201907002246. Archived from the original on December 30, 2016.
- Pol, D.; Rauhut, O.W.M. (2012). "A Middle Jurassic abelisaurid from Patagonia and the early diversification of theropod dinosaurs". Proceedings of the Royal Society B. 279 (1741): 3170–5. doi:10.1098/rspb.2012.0660. PMC 3385738. PMID 22628475.
- Farke, A.A.; Sertich, J.J.W (2013). "An Abelisauroid Theropod Dinosaur from the Turonian of Madagascar". PLOS One. 8 (4): e62047. Bibcode:2013PLoSO...862047F. doi:10.1371/journal.pone.0062047. PMC 3630149. PMID 23637961.
- Ezcurra, M.D.; Agnolin, F.L.; Novas, F.E. (2010). "An abelisauroid dinosaur with a non-atrophied manus from the Late Cretaceous Pari Aike Formation of southern Patagonia". Zootaxa. 2450 (2450): 1. doi:10.11646/zootaxa.2450.1.1.
- Rauhut, O.W.M.; Carrano, M.T. (2016). "The theropod dinosaur Elaphrosaurus bambergi Janensch, 1920, from the Late Jurassic of Tendaguru, Tanzania". Zoological Journal of the Linnean Society. 178 (3): 546–610. doi:10.1111/zoj.12425.
- Carrano, M.T.; Benson, R.B.J.; Sampson, S.D. (2012). "The phylogeny of Tetanurae (Dinosauria: Theropoda)". Journal of Systematic Palaeontology. 10 (2): 211–300. doi:10.1080/14772019.2011.630927.
- Langer, Max Cardoso; Martins, Neurides de Oliveira; Manzig, Paulo César; Ferreira, Gabriel de Souza; Marsola, Júlio César de Almeida; Fortes, Edison; Lima, Rosana; Sant’ana, Lucas Cesar Frediani; Vidal, Luciano da Silva; Lorençato, Rosangela Honório da Silva; Ezcurra, Martín Daniel (June 26, 2019). "A new desert-dwelling dinosaur (Theropoda, Noasaurinae) from the Cretaceous of south Brazil". Scientific Reports. 9 (1): 9379. Bibcode:2019NatSR...9.9379L. doi:10.1038/s41598-019-45306-9. ISSN 2045-2322. PMC 6594977. PMID 31243312.
- "New dinosaur gives bird wing clue". BBC News. June 17, 2009. Retrieved April 19, 2020.
- Xu, X.; Mackem, S. (2013). "Tracing the evolution of avian wing digits". Current Biology. 23 (12): 538–544. doi:10.1016/j.cub.2013.04.071. PMID 23787052.
- Larsson, H.C.E.; Wagner, G.P. (2002). "Pentadactyl ground state of the avian wing". Journal of Experimental Zoology. 294 (2): 146–151. doi:10.1002/jez.10153. PMID 12210115.
- Feduccia, A. (2002). "Birds are dinosaurs: simple answer to a complex problem". The Auk. 119 (4): 1187–1201. doi:10.1642/0004-8038(2002)119[1187:BADSAT]2.0.CO;2.
- Thulborn, R.A.; Hamley, T.L. (1982). "The reptilian relationships of Archaeopteryx". Australian Journal of Zoology. 30 (4): 611–634. doi:10.1071/ZO9820611.
- Kaplan, M. (2009). "Dinosaur's digits show how birds got wings". Nature. doi:10.1038/news.2009.577.
- Wagner, G.P.; Gauthier, J.A. (1999). "1,2,3 = 2,3,4: A solution to the problem of the homology of the digits in the avian hand". Proceedings of the National Academy of Sciences. 96 (9): 5111–5116. Bibcode:1999PNAS...96.5111W. doi:10.1073/pnas.96.9.5111. PMC 21825. PMID 10220427.
- Vargas, A.O.; Wagner, G.P. (2009). "Frame-shifts of digit identity in bird evolution and Cyclopamine-treated wings". Evolution & Development. 11 (2): 163–169. CiteSeerX 10.1.1.595.7937. doi:10.1111/j.1525-142X.2009.00317.x. PMID 19245548.
- Bever, G.S.; Gauthier, J.A.; Wagner, G.P. (2011). "Finding the frame shift: digit loss, developmental variability, and the origin of the avian hand". Evolution & Development. 13 (3): 269–279. doi:10.1111/j.1525-142X.2011.00478.x. PMID 21535465.
- Young, R.L.; Bever, G.S.; Wang, Z.; Wagner, G.P. (2011). "Identity of the avian wing digits: Problems resolved and unsolved". Developmental Dynamics. 240 (5): 1042–1053. doi:10.1002/dvdy.22595. PMID 21412936.
- Vargas, A.O.; Wagner, G.P.; Gauthier, J.A. (2009). "Limusaurus and bird digit identity". Nature Precedings. doi:10.1038/npre.2009.3828.1. Archived from the original on December 23, 2016. Retrieved December 22, 2016.
- Čapek, D.; Metscher, B.D.; Muller, G.B. (2014). "Thumbs down: a molecular‐morphogenetic approach to avian digit homology". Journal of Experimental Zoology. 322 (1): 1–12. doi:10.1002/jez.b.22545. PMID 24323741.
- Xu, X.; Clark, J.; Choiniere, J.; Hone, D.; Sullivan, C. (2011). "Reply to "Limusaurus and bird digit identity"". Nature Precedings. doi:10.1038/npre.2011.6375.1. Archived from the original on December 30, 2016. Retrieved December 29, 2016.
- Carrano, M.T.; Choiniere, J. (2016). "New information on the forearm and manus of Ceratosaurus nasicornis Marsh, 1884 (Dinosauria, Theropoda), with implications for theropod forelimb evolution". Journal of Vertebrate Paleontology. 36 (2): e1054497. doi:10.1080/02724634.2015.1054497.
- Louchart, A.; Viriot, L. (2011). "From snout to beak: the loss of teeth in birds". Trends in Ecology and Evolution. 26 (12): 663–673. doi:10.1016/j.tree.2011.09.004. PMID 21978465.
- Wang, Shuo; Stiegler, Josef; Wu, Ping; Chuong, Cheng-Ming; Hu, Dongyu; Balanoff, Amy; Zhou, Yachun; Xu, Xing (2017). "Heterochronic truncation of odontogenesis in theropod dinosaurs provides insight into the macroevolution of avian beaks". Proceedings of the National Academy of Sciences of the United States of America. 114 (41): 10930–10935. doi:10.1073/pnas.1708023114. PMC 5642708. PMID 28973883.
- Buchtova, M.; Zahradnicek, O.; Balkova, S.; Tucker, A.S. (2013). "Odontogenesis in the Veiled Chameleon (Chamaeleo calyptratus)". Archives of Oral Biology. 58 (2): 118–133. doi:10.1016/j.archoralbio.2012.10.019. PMID 23200300. Archived from the original on April 20, 2013. Retrieved December 22, 2016.
- Chen, YP; Zhang, Y.; Jiang, T.-X.; Barlow, A.J.; St. Amand, T.R.; Hu, Y.; Heaney, S.; Francis-West, P.; Chuong, C.-M.; Maas, R. (2000). "Conservation of early odontogenic signaling pathways in Aves". Proceedings of the National Academy of Sciences of the United States of America. 97 (18): 10044–10049. Bibcode:2000PNAS...9710044C. doi:10.1073/pnas.160245097. PMC 27667. PMID 10954731.
- Zanno, L.E.; Makovicky, P.J. (2011). "Herbivorous ecomorphology and specialization patterns in theropod dinosaur evolution". Proceedings of the National Academy of Sciences of the United States of America. 108 (1): 232–237. Bibcode:2011PNAS..108..232Z. doi:10.1073/pnas.1011924108. PMC 3017133. PMID 21173263.
- Delcourt, R. (2018). "Ceratosaur palaeobiology: new insights on evolution and ecology of the southern rulers". Scientific Reports. 8 (1). doi:10.1038/s41598-018-28154-x.
- Eberth, D.A.; Brinkman, D.B.; Chen, P.-J.; Yuan, F.-T.; Wu, S.Z.; Li, G.; Cheng, X.-S. (2001). "Sequence stratigraphy, paleoclimate patterns, and vertebrate fossil preservation in Jurassic–Cretaceous strata of the Junggar Basin, Xinjiang Autonomous Region, People's Republic of China". Canadian Journal of Earth Sciences. 38 (12): 1627–1644. Bibcode:2001CaJES..38.1627E. doi:10.1139/e01-067.
- Tutken, T.; Pfretzschner, H.-U.; Vennemann, T.W.; Sun, G.; Wang, Y.D. (2004). "Paleobiology and skeletochronology of Jurassic dinosaurs: implications from the histology and oxygen isotope compositions of bones". Palaeogeography, Palaeoclimatology, Palaeoecology. 206 (3): 217–238. Bibcode:2004PPP...206..217T. doi:10.1016/j.palaeo.2004.01.005.
- Hinz, J.K.; Smith, I.; Pfretzschner, H.-U.; Wings, O.; Sun, G. (2010). "A high-resolution three-dimensional reconstruction of a fossil forest (Upper Jurassic Shishugou Formation, Junggar Basin, Northwest China)". Palaeobiodiversity and Palaeoenvironments. 90 (3): 215–240. doi:10.1007/s12549-010-0036-y.
- He, Y.-M.; Clark, J.M.; Xu, X. (2013). "A large theropod metatarsal from the upper part of Jurassic Shishugou Formation in Junggar Basin, Xinjiang, China" (PDF). Vertebrata PalAsiatica. 51 (1): 29–42. Archived (PDF) from the original on April 2, 2014. Retrieved December 30, 2016.
- Handwerk, B. (January 20, 2010). "Dinosaur "Death Pits" Created by Giant's Footprints?". National Geographic News. Retrieved June 26, 2020.