DPANN
| DPANN | |
|---|---|
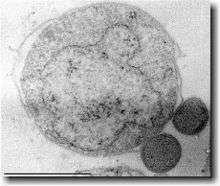 | |
| Nanoarchaeum equitans (the small symbionts on the right) | |
| Scientific classification | |
| Domain: | Archaea |
| Superphylum: | DPANN group |
| Phyla | |
DPANN (Diapherotrites, Parvarchaeota, Aenigmarchaeota, Nanoarchaeota, Nanohaloarchaea) is a superphylum of Extremophile Archaea.[1]
It was proposed in 2013 by Rinke et al.[2]
DPANN is an acronym formed by the initials of the first five groups discovered, Diapherotrites, Parvarchaeota, Aenigmarchaeota, Nanoarchaeota and Nanohaloarchaeota. Later Woesearchaeota and Pacearchaeota were discovered and proposed within the DPANN superphylum.[1] In 2017, another phylum Altiarchaeota was placed into this superphylum.[3]
The DPANN groups together different phyla with a variety of environmental distribution and metabolism, ranging from symbiotic and thermophilic forms such as Nanoarchaeota, acidophiles like Parvarchaeota and non-extremophiles like Aenigmarchaeota and Diapherotrites.
Classification
- Diapherotrites. Found by phylogenetic analysis of the genomes recovered from the groundwater filtration of a gold mine abandoned in the USA.[4][5]
- Parvarchaeota and Micrarchaeota. Discovered in 2006 in strongly acidic sewage from a US mine.[6][7][8] They are of very small size and provisionally called ARMAN (Archaeal Richmond Mine Acidophilic Nanoorganisms).
- Woesearchaeota and Pacearchaeota. They have been identified both in sediments and in surface waters of aquifers and lakes, abounding especially in saline conditions.[9][10]
- Aenigmarchaeota. Found in wastewater from mines and in sediments from hot springs.[11]
- Nanohaloarchaeota. Distributed in environments with high salinity.[12]
- Nanoarchaeota. They were the first discovered (in 2002) in a hydrothermal source next to the coast of Iceland. They live as symbionts of other archaea.[13][14][15]
References
- 1 2 Cindy J. Castelle, Kelly C. Wrighton, ..., Kenneth H. Williams, Jillian F. Banfield (16 March 2015). "Genomic Expansion of Domain Archaea Highlights Roles for Organisms from New Phyla in Anaerobic Carbon Cycling" (PDF). Current Biology. Retrieved 14 January 2017.
- ↑ Rinke, C., Schwientek, P., Sczyrba, A., Ivanova, N. N., Anderson, I. J., Cheng, J. F., ... & Dodsworth, J. A. (2013). Insights into the phylogeny and coding potential of microbial dark matter. Nature, 499(7459), 431-437.
- ↑ Anja Spang, Eva F. Caceres, Thijs J. G. Ettema (11 Aug 2017). "Genomic exploration of the diversity, ecology, and evolution of the archaeal domain of life". Science.
- ↑ Genomes Online Database
- ↑ Comolli LR, Baker BJ, Downing KH, Siegerist CE, Banfield JF (2009) Three-dimensional analysis of the structure and ecology of a novel, ultra-small archaeon. ISME J 3:159–167.
- ↑ Baker, B. J., Tyson, G. W., Webb, R. I., et al.(2006) “Lineages of acidophilic archaea revealed by community genomic analysis,” Science, 314(5807):1933–1935.
- ↑ Murakami, S., Fujishima, K., Tomita, M., Kanai, A.(2012) Metatranscriptomic analysis of microbes in an ocean-front deep subsurface hot spring reveals novel small RNAs and type-specific tRNA degradation. Applied and Environmental Microbiology 78(4):1015-22.
- ↑ B. J. Baker, L. R. Comolli, G. J. Dick, L. J. Hauser, D. Hyatt, B. D. Dill, M. L. Land, N. C. VerBerkmoes, R. L. Hettich, J. F. Banfield. Enigmatic, ultrasmall, uncultivated Archaea. Proceedings of the National Academy of Sciences (2010).
- ↑ Castelle, C. J., Wrighton, K. C., Thomas, B. C., Hug, L. A., Brown, C. T., Wilkins, M. J., ... & Taylor, R. C. (2015). Genomic expansion of domain archaea highlights roles for organisms from new phyla in anaerobic carbon cycling. Current Biology, 25(6), 690-701.
- ↑ Ortiz‐Alvarez, R., & Casamayor, E. O. (2016). High occurrence of Pacearchaeota and Woesearchaeota (Archaea superphylum DPANN) in the surface waters of oligotrophic high‐altitude lakes. Environmental microbiology reports.
- ↑ Takai, K., Moser, D. P., DeFlaun, M., Onstott, T. C. & Fredrickson, J. K. Archaeal diversity in waters fromdeep south african gold mines. Appl. Environ. Microbiol. 67, 5750–5760 (2001).
- ↑ Narasingarao, P. et al. (2012). "De novo metagenomic assembly reveals abundant novel major lineage of Archaea in hypersaline microbial communities". ISME J. 6 (1): 81–93.
- ↑ Waters, E., Hohn, M. J., Ahel, I., Graham, D. E., Adams, M. D., Barnstead, M., Beeson, K. Y., Bibbs, L., Bolanos, R., Keller, M., Kretz, K., Lin, X., Mathur, E., Ni, J., Podar, M., Richardson, T., Sutton, G. G., Simon, M., Soll, D., Stetter, K. O., Short, J. M., Noordewier, M. (2003).
- ↑ “The genome of Nanoarchaeum equitans: insights into early archaeal evolution and derived parasitism”. Proc Natl Acad Sci U S A 100 (22): 12984–8.
- ↑ Podar, M., Makarova, K. S., Graham, D. E., Wolf, Y. I., Koonin, E. V., Reysenbach, A. L. (2013). Insights into archaeal evolution and symbiosis from the genomes of a nanoarchaeon and its inferred crenarchaeal host from Obsidian Pool, Yellowstone National Park. Biol. Direct 8:9
External links
