Megalodon
| Megalodon | |
|---|---|
 | |
| Model of megalodon jaws at the American Museum of Natural History | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Chondrichthyes |
| Order: | Lamniformes |
| Family: | †Otodontidae |
| Genus: | †Carcharocles |
| Species: | †C. megalodon |
| Binomial name | |
| Carcharocles megalodon | |
| Synonyms[2][3][4][5] | |
|
List of synonyms
| |
Megalodon (Carcharocles megalodon), meaning "big tooth", is an extinct species of shark that lived approximately 23 to 2.6 million years ago (mya), during the Early Miocene to the end of the Pliocene. It was formerly thought to belong to the family Lamnidae, making it closely related to the great white shark (Carcharodon carcharias). However presently there is near unanimous consensus that it belongs to the extinct family Otodontidae, which diverged from the ancestry of the great white shark during the Early Cretaceous. Its genus placement is still debated, authors placing it in either Carcharocles, Megaselachus, Otodus, or Procarcharodon.
Scientists suggest that megalodon looked like a stockier version of the great white shark, though it may have looked similar to the basking shark (Cetorhinus maximus) or the sand tiger shark (Carcharias taurus). Regarded as one of the largest and most powerful predators to have ever lived, fossil remains of megalodon suggest that this giant shark reached a length of 18 meters (59 ft).[6] Their large jaws could exert a bite force of up to 110,000 to 180,000 newtons (24,000 - 41,000 lbf). Their teeth were thick and robust, built for grabbing prey and breaking bone.
Megalodon probably had a major impact on the structure of marine communities. The fossil record indicates that it had a cosmopolitan distribution. It probably targeted large prey, such as whales, seals, and giant turtles. Juveniles inhabited warm coastal waters where they would feed on fish and small whales. Unlike the great white, which attacks prey from the soft underside, megalodon probably used its strong jaws to break through the chest cavity and puncture the heart and lungs of its prey.
The animal faced competition from whale-eating cetaceans, such as Livyatan and ancient killer whales (Orcinus citoniensis), which likely contributed to its extinction. As it preferred warmer waters, it is thought that oceanic cooling associated with the onset of the ice ages, coupled with the lowering of sea levels and resulting loss of suitable nursery areas, may have also contributed to its decline. A reduction in the diversity of baleen whales and a shift in their distribution toward polar regions may have reduced megalodon's primary food source. The extinction of the shark appeared to affect other animals; for example, the size of baleen whales increased significantly after the shark had disappeared.
Taxonomy
Naming

According to Renaissance accounts, gigantic, triangular fossil teeth often found embedded in rocky formations were once believed to be the petrified tongues, or glossopetrae, of dragons and snakes. This interpretation was corrected in 1667 by Danish naturalist Nicolas Steno, who recognized them as shark teeth, and famously produced a depiction of a shark's head bearing such teeth. He described his findings in the book The Head of a Shark Dissected, which also contained an illustration of a megalodon tooth.[7][8][9]
Swiss naturalist Louis Agassiz gave this shark its initial scientific name, Carcharodon megalodon, in his 1843 work Recherches sur les poissons fossiles, based on tooth remains.[1][10] English paleontologist Edward Charlesworth in his 1837 paper used the name Carcharias megalodon, while citing Agassiz as the author, indicating that Agassiz described the species prior to 1843. English paleontologist Charles Davies Sherborn in 1928 listed an 1835 series of articles by Agassiz as the first scientific description of the shark.[11] The specific name megalodon translates to "big tooth", from Ancient Greek: μέγας, translit. (megas), lit. 'big, mighty' and οδόντος (odontús), "tooth".[12][13] The teeth of megalodon are morphologically similar to those of the great white shark (Carcharodon carcharias), and on the basis of this observation, Agassiz assigned megalodon to the genus Carcharodon.[10] Though “megalodon” is an informal name for the shark, it is also often informally dubbed the "giant white shark",[14] the "megatooth shark", the "big tooth shark", or "Meg".[15]:4
Evolution
|
| ||||||||||||||||||||||||
| Relationship between megalodon and other sharks, including the great white shark (Carcharodon carcharias)[16] |
While the earliest megalodon remains were reported from the Late Oligocene dated to around 28 million years ago (mya),[17][18] competing figures still exist as to when it evolved, such as 16 mya and 23 mya.[19] It is believed that megalodon became extinct around the end of the Pliocene, probably about 2.6 mya;[19][20] reported Pleistocene megalodon teeth, younger than 2.6 million years old, are considered to be unreliable claims.[20]
Megalodon is now considered to be a member of the family Otodontidae, genus Carcharocles, as opposed to its previous classification into Lamnidae, genus Carcharodon.[19][20][21][22] Megalodon's classification into Carcharodon was due to dental similarity with the great white shark, but most authors currently believe that this is due to convergent evolution. In this model, the great white shark is more closely related to the shark Isurus hastalis than to megalodon, as evidenced by more similar dentition in those two sharks; megalodon teeth have much finer serrations than great white shark teeth. The great white shark is more closely related to the mako shark (Isurus spp.), with a common ancestor around 4 mya.[10][16] Proponents of the former model, wherein megalodon and the great white shark are more closely related, argue that the differences between their dentition are minute and obscure.[23]:23–25

The genus Carcharocles currently contains four species: C. auriculatus, C. angustidens, C. chubutensis, and C. megalodon.[15]:30–31 The genus was proposed by D. S. Jordan and H. Hannibal in 1923 to contain C. auriculatus. In the 1980s, megalodon was assigned to Carcharocles.[10][15]:30 Before this, in 1960, the genus Procarcharodon was erected by French ichthyologist Edgard Casier, which included those four sharks and was considered separate from the great white shark. It is now considered a junior synonym of Carcharocles.[15]:30 The genus Palaeocarcharodon was erected alongside Procarcharodon to represent the beginning of the lineage, and, in the model wherein megalodon and the great white shark are closely related, their last common ancestor. It is believed to be an evolutionary dead-end and unrelated to the Carcharocles sharks by authors who reject that model.[23]:70

Another model of the evolution of this genus, also proposed by Casier in 1960, is that the direct ancestor of the Carcharocles is the shark Otodus obliquus, which lived from 60 mya to 13 mya during the Paleocene and Miocene epochs.[16][24] In this model, O. obliquus evolved into O. aksuaticus, which evolved into C. auriculatus, and then into C. angustidens, and then into C. chubutensis, and then finally into C. megalodon. The evolution of this lineage is characterized by the increase of serrations, the widening of the crown, the development of a more triangular shape, and the disappearance of the lateral cusps.[15]:28–31[24] The genus Otodus is ultimately derived from Cretolamna, a shark from the Cretaceous period.[21][25]
Another model of the evolution of Carcharocles, proposed in 2001 by paleontologist Michael Benton, is that the three other species are actually a single species of shark that gradually changed over time between the Paleocene and the Pliocene, making it a chronospecies.[15]:17[18][26] Some authors suggest that C. auriculatus, C. angustidens, and C. chubutensis should be classified as a single species in the genus Otodus, leaving C. megalodon the sole member of Carcharocles.[18][27]
The genus Carcharocles may be invalid, and the shark may actually belong in the genus Otodus, making it Otodus megalodon.[4] A 1974 study on Paleogene sharks by Henri Cappetta erected the subgenus Megaselachus, classifying the shark as Otodus (Megaselachus) megalodon, along with O. (M.) chubutensis. A 2006 review of Chondrichthyes elevated Megaselachus to genus, and classified the sharks as Megaselachus megalodon and M. chubutensis.[4] The discovery of fossils assigned to the genus Megalolamna in 2016 led to a re-evaluation of Otodus, which concluded that it is paraphyletic, that is, it consists of a last common ancestor but it does not include all of its descendants. The inclusion of the Carcharocles sharks in Otodus would make it monophyletic, with the sister clade being Megalolamna.[21] There was one apparent description of the shark in 1881 classifying it as Selache manzonii.[28]
Biology
Appearance

One interpretation on how megalodon appeared was that it was a robust-looking shark, and may have had a similar build to the great white shark. The jaws may have been blunter and wider than the great white, and the fins would have also been similar in shape, though thicker due to its size. It may have had a pig-eyed appearance, in that it had small, deep-set eyes.[23]:64–65
Another interpretation is that megalodon bore a similarity to the whale shark (Rhincodon typus) or the basking shark (Cetorhinus maximus). The tail fin would have been crescent-shaped, the anal fin and second dorsal fin would have been small, and there would have been a caudal keel present on either side of the tail fin (on the caudal peduncle). This build is common in other large aquatic animals, such as whales, tuna, and other sharks, in order to reduce drag while swimming. The head shape can vary between species as most of the drag-reducing adaptations are toward the tail-end of the animal.[15]:35–36

Since Carcharocles is derived from Otodus, and the two had teeth that bear a close similarity to those of the sand tiger shark (Carcharias taurus), megalodon may have had a build more similar to the sand tiger shark than to other sharks. This is unlikely since the sand tiger shark is a carangiform swimmer which require faster movement of the tail for propulsion through the water than the great white shark, a thunniform swimmer.[15]:35–36[29]
Size
Statistics
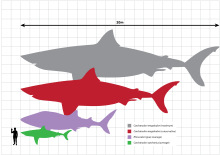
Due to fragmentary remains, there have been many contradictory size estimates for megalodon, as they can only be drawn from fossil teeth and vertebrae.[15]:87[30] Also because of this, the great white shark is the basis of its reconstruction and size estimation,[23]:57 as it is regarded as the best analogue to megalodon.[27] Various size estimates exist for megalodon; in 1973, Hawaiian ichthyologist John E. Randall estimated that the maximum length attained by C. megalodon was about 13 meters (43 ft),[31] while in the 1990s, marine biologists Patrick J. Schembri and Stephen Papson opined that C. megalodon may have approached a maximum of around 24 to 25 meters (79 to 82 ft) in total length;[32][33] Gottfried and colleagues asserted that C. megalodon could have reached a maximum of 20.3 meters (67 ft) in total length.[34][23][35] Nowadays, the commonly acknowledged maximum total length of C. megalodon is about 18 meters (59 ft), with the average size being 10.5 meters (34 ft),[19][20][22] compared to the maximum recorded sizes of the great white shark at 6.1 meters (20 ft) and the whale shark (the largest extant fish) at 12.65 m (42 ft).[36][37][38][39] It is possible that different populations of megalodon around the globe had different body sizes and behaviors due to different ecological pressures.[22] If it did attain a size of over 16 meters (52 ft), it would have been the largest known fish that has ever lived, surpassing the Jurassic fish Leedsichthys.[40]
Mature male megalodon may have had a body mass of 12.6 to 33.9 metric tons (13.9 to 37.4 short tons), and mature females may have been 27.4 to 59.4 metric tons (30.2 to 65.5 short tons), given that males could range in length from 10.5 to 14.3 meters (34 to 47 ft) and females 13.3 to 17 meters (44 to 56 ft).[23]:61[34] A 2015 study linking shark size and typical swimming speed estimated that megalodon would have typically swum at 18 kilometers per hour (11 mph), given that its body mass was typically 48 metric tons (53 short tons), which is consistent with other aquatic creatures of its size, such as the fin whale which typically cruises at speeds of 14.5 to 21.5 km/h (9.0 to 13.4 mph).[41]
Its large size may have been due to climatic factors and the abundance of large prey items, and it may have also been influenced by the evolution of regional endothermy (mesothermy) which would have increased its metabolic rate and swimming speed. Since the otodontid sharks are considered to have been ectotherms, and megalodon was a close relative to them, megalodon may have also been ectothermic. Contrary to this, the largest contemporary ectothermic sharks, such as the whale shark, are filter feeders, implying some metabolic constraints with a predatory lifestyle. That is to say, it is unlikely that megalodon was ectothermic.[42]
Estimations
Gordon Hubbell from Gainesville, Florida, possesses an upper anterior megalodon tooth whose maximum height is 18.4 centimeters (7.25 in), one of the largest known tooth specimens from the shark.[43] In addition, a 2.7-by-3.4-meter (9 by 11 ft) megalodon jaw reconstruction developed by fossil hunter Vito Bertucci contains a tooth whose maximum height is reportedly over 18 centimeters (7 in).[44]
The first attempt to reconstruct the jaw of megalodon was made by Bashford Dean in 1909, displayed at the American Museum of Natural History. From the dimensions of this jaw reconstruction, it was hypothesized that megalodon could have approached 30 meters (98 ft) in length. Dean had overestimated the size of the cartilage on both jaws, causing it to be too tall.[40][45]
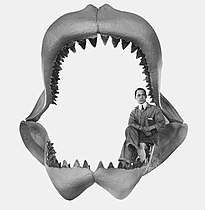

John E. Randall, an ichthyologist, used the enamel height (the vertical distance of the blade from the base of the enamel portion of the tooth to its tip) to measure the length of the shark, yielding a maximum length of about 13 meters (43 ft).[31] Tooth enamel height does not necessarily increase in proportion to the animal's total length.[23]:99
Shark researchers Michael D. Gottfried, Leonard Compagno, and S. Curtis Bowman proposed a linear relationship between a shark's total length and the height of the largest upper anterior tooth. The proposed relationship is: total length in meters = − (0.096) × [UA maximum height (mm)]-(0.22).[18][23]:60 They had estimated the average height, based on the slant height of the largest tooth discovered, for large female megalodon to be 15.6 meters (51 ft), though larger teeth may exist.[23]:55–60
In 2002, shark researcher Clifford Jeremiah proposed that total length was proportional to the root width of an upper anterior tooth. He claimed that for every 1 centimeter (0.39 in) of root width, there are approximately 1.4 meters (4.6 ft) of shark length. Jeremiah pointed out that the jaw perimeter of a shark is directly proportional to its total length, with the width of the roots of the largest teeth being a tool for estimating jaw perimeter. The largest tooth in Jeremiah's possession had a root width of about 12 centimeters (4.7 in), which yielded 16.5 meters (54 ft) in total length.[15]:88
In 2002, paleontologist Kenshu Shimada of DePaul University proposed a linear relationship between tooth crown height and total length after conducting anatomical analysis of several specimens, allowing any sized tooth to be used. Shimada stated that the previously proposed methods were based on a less-reliable evaluation of the dental homology between megalodon and the great white shark, and that the growth rate between the crown and root is not isometric, which he considered in his model. Using this model, the upper anterior tooth possessed by Gottfried and colleagues corresponded to a total length of 15 meters (49 ft).[46] Among the specimens found in the Gatún Formation of Panama, other shark researchers used this method and calculated a maximum length of 16.8 meters (55 ft) for a specimen,[27] and for another a total length of 17.9 meters (59 ft). This result appears to be an error within the matrix, and the length of this individual is actually 19.6 meters (64 ft).[47]
Teeth and bite force

The most common fossils of megalodon are its teeth. Diagnostic characteristics include a triangular shape, robust structure, large size, fine serrations, a lack of lateral denticles, and a visible V-shaped neck (where the root meets the crown).[23]:55[27] The tooth met the jaw at a steep angle, similar to the great white shark. The tooth was anchored by connective tissue fibers, and the roughness of the base may have added to mechanical strength.[48] The lingual side of the tooth, the part facing the tongue, was convex; and the labial side, the other side of the tooth, was slightly convex or flat. The anterior teeth were almost perpendicular to the jaw and symmetrical, whereas the posterior teeth were slanted and asymmetrical.[49]
Megalodon teeth can measure over 180 millimeters (7.1 in) in slant height (diagonal length) and are the largest of any known shark species.[15]:33 In 1989, a nearly complete set of megalodon teeth was discovered in Saitama, Japan. Another nearly complete associated megalodon dentition was excavated from the Yorktown Formations in the United States, and served as the basis of a jaw reconstruction of megalodon at the National Museum of Natural History (USNM). Based on these discoveries, an artificial dental formula was put together for megalodon in 1996.[23]:55[50]
The dental formula of megalodon is: 2.1.7.43.0.8.4. As evident from the formula, megalodon had four kinds of teeth in its jaws: anterior, intermediate, lateral, and posterior. Megalodon's intermediate tooth technically appears to be an upper anterior and is termed as "A3" because it is fairly symmetrical and does not point mesially (side of the tooth toward the midline of the jaws where the left and right jaws meet). Megalodon had a very robust dentition,[23]:20–21 and had over 250 teeth in its jaws, spanning 5 rows.[15]:iv It is possible that large megalodon individuals had jaws spanning roughly 2 meters (6.6 ft) across.[15]:129 The teeth were also serrated, which would have improved efficiency in cutting through flesh or bone.[10][15]:1 The shark may have been able to open its mouth to a 75° angle, though a reconstruction at the USNM approximates a 100° angle.[23]:63

In 2008, a team of scientists led by S. Wroe conducted an experiment to determine the bite force of the great white shark, using a 2.5-meter (8.2 ft) long specimen, and then isometrically scaling the results for its maximum confirmed size and the conservative minimum and maximum body mass of megalodon. They placed the bite force of the latter between 110,000 to 180,000 newtons (24,000 to 41,000 lbf) in a posterior bite, compared to the 18,000 N (4,100 lbf) bite force for the largest confirmed great white shark, and 7,400 N (1,700 lbf) for the placoderm fish Dunkleosteus. In addition, Wroe and colleagues pointed out that sharks shake sideways while feeding, amplifying the force generated, which would probably have caused the total force experienced by prey to be higher than the estimate.[34][51]
Internal anatomy

Megalodon is represented in the fossil record by teeth, vertebral centra, and coprolites.[23]:57 As with all sharks, the skeleton of megalodon was formed of cartilage rather than bone; consequently most fossil specimens are poorly preserved.[52] To support its large dentition, the jaws of megalodon would have been more massive, stouter, and more strongly developed than those of the great white, which possesses a comparatively gracile dentition. Its chondrocranium, the cartilaginous skull, would have had a blockier and more robust appearance than that of the great white. Its fins were proportional to its larger size.[23]:64–65
Some fossil vertebrae have been found. The most notable example is a partially preserved vertebral column of a single specimen, excavated in the Antwerp Basin, Belgium, in 1926. It comprises 150 vertebral centra, with the centra ranging from 55 millimeters (2.2 in) to 155 millimeters (6 in) in diameter. The shark's vertebrae may have gotten much bigger, and scrutiny of the specimen revealed that it had a higher vertebral count than specimens of any known shark, possibly over 200 centra; only the great white approached it.[23]:63–65 Another partially preserved vertebral column of a megalodon was excavated from the Gram Formation in Denmark in 1983, which comprises 20 vertebral centra, with the centra ranging from 100 millimeters (4 in) to 230 millimeters (9 in) in diameter.[48]
.jpg)
The coprolite remains of megalodon are spiral-shaped, indicating that the shark may have had a spiral valve, a corkscrew-shaped portion of the lower intestines, similar to extant lamniform sharks. Miocene coprolite remains were discovered in Beaufort County, South Carolina, with one measuring 14 cm (5.5 in).[53]
Gottfried and colleagues reconstructed the entire skeleton of megalodon, which was later put on display at the Calvert Marine Museum in the United States and the Iziko South African Museum.[23]:56[24] This reconstruction is 11.3 meters (37 ft) long and represents a mature male,[23]:61 based on the ontogenetic changes a great white shark experiences over the course of its life.[23]:65
Paleobiology
Range and habitat
Megalodon had a cosmopolitan distribution;[19][47] its fossils have been excavated from many parts of the world, including Europe, Africa, the Americas, and Australia.[23]:67[54] It most commonly occurred in subtropical to temperate latitudes.[19][23]:78 It has been found at latitudes up to 55° N; its inferred tolerated temperature range was 1–24 °C (34–75 °F). It arguably had the capacity to endure such low temperatures due to mesothermy, the physiological capability of large sharks to conserve metabolic heat by maintaining a higher body temperature than the surrounding water.[19]
Megalodon inhabited a wide range of marine environments (i.e., shallow coastal waters, areas of coastal upwelling, swampy coastal lagoons, sandy littorals, and offshore deep water environments), and exhibited a transient lifestyle. Adult megalodon were not abundant in shallow water environments, and mostly inhabited offshore areas. Megalodon may have moved between coastal and oceanic waters, particularly in different stages of its life cycle.[15]:33[55] Megalodon teeth have been excavated from regions far away from continental lands, such as the Marianas Trench in the Pacific Ocean.[15]:iv
Fossil remains show a trend for specimens to be larger on average in the southern hemisphere than in the northern, with mean lengths of 11.6 and 9.6 meters (38 and 31 ft), respectively; and also larger in the Pacific than the Atlantic, with mean lengths of 10.9 and 9.5 meters (36 and 31 ft) respectively. They do not suggest any trend of changing body size with absolute latitude, or of change in size over time (although the Carcharocles lineage in general is thought to display a trend of increasing size over time). The overall modal length has been estimated at 10.5 meters (34 ft), with the length distribution skewed towards larger individuals, suggesting an ecological or competitive advantage for larger body size.[22]
Locations of fossils
Megalodon had a global distribution and fossils of the shark have been found in many places around the world, bordering all oceans of the Neogene.[56]
Prey relationships

Though sharks are generally opportunistic feeders, megalodon's great size, high-speed swimming capability, and powerful jaws, coupled with an impressive feeding apparatus, made it an apex predator capable of consuming a broad spectrum of animals. It was probably one of the most powerful predators to have existed.[23]:71–75[34] A study focusing on calcium isotopes of extinct and extant elasmobranch sharks and rays revealed that megalodon fed at a higher trophic level than the contemporaneous great white shark, that is to say it was higher up in the food chain.[57]
Fossil evidence indicates that megalodon preyed upon many cetacean species, such as dolphins, small whales, cetotheres, squalodontids, sperm whales, bowhead whales, and rorquals.[40][58][59] In addition to this, they also targeted seals, sirenians, and large sea turtles.[55] The shark was an opportunist and piscivorous, and it would have also gone after smaller fish and other sharks.[40] Many whale bones have been found with deep gashes most likely made by their teeth.[23]:75 Various excavations have revealed megalodon teeth lying close to the chewed remains of whales,[23]:75[24] and sometimes in direct association with them.[14]
The feeding ecology of megalodon appears to have varied with age and between sites, like the modern great white. It is plausible that the adult megalodon population off the coast of Peru targeted primarily cetothere whales 2.5 to 7 meters (8.2 to 23 ft) in length and other prey smaller than itself, rather than large whales in the same size class as themselves.[58] Meanwhile, juveniles likely had a diet that consisted more of fish.[27][60]
Competition

Megalodon faced a highly competitive environment.[61] Its position at the top of the food chain[62] probably had a significant impact on the structuring of marine communities.[61][63] Fossil evidence indicates a correlation between megalodon and the emergence and diversification of cetaceans and other marine mammals.[23]:78[61] Juvenile megalodon preferred habitats where small cetaceans were abundant, and adult megalodon preferred habitats where large cetaceans were abundant. Such preferences may have developed shortly after they appeared in the Oligocene.[23]:74–75
Megalodon were contemporaneous with whale-eating odontocetes (particularly macroraptorial sperm whales and squalodontids), which were also probably among the era's apex predators, and provided competition.[61] In response to competition from megalodon, whale-eating odontocetes may have evolved defensive adaptations; some species, such as the killer whale (Orcinus citoniensis), became pack predators,[24][64] and some attained gigantic sizes, such as Livyatan, which grew from 13.5 to 17.5 meters (44 to 57 ft). By the Late Miocene, around 11 mya, macroraptorials experienced a significant decline in abundance and diversity. Other hyper-predatory cetaceans began to emerge during the Pliocene to fill this ecological void, such as the killer whale.[61][65]
Fossil evidence indicates that other notable species of sharks, such as the great white shark, responded to competitive pressure from megalodon by avoiding regions it inhabited by mainly inhabiting the cooler waters of the time. In areas where their ranges seemed to overlap, such as in Pliocene Baja California, it is possible that megalodon and the great white shark occupied the area at different times of the year while following different migratory prey.[23]:77[66] Megalodon probably also had a tendency for cannibalism, much like contemporary sharks.[67]
Feeding strategies

Sharks often employ complex hunting strategies to engage large prey animals. Great white shark hunting strategies may be similar to how megalodon hunted its large prey.[68] Megalodon bite marks on whale fossils suggests that it employed different hunting strategies against large prey than the great white shark.[40]
One particular specimen – the remains of a 9-meter (30 ft) long prehistoric baleen whale (of an unknown Miocene taxon) – provided the first opportunity to quantitatively analyze its attack behavior. Unlike great whites which target the underbelly of their prey, megalodon probably targeted the heart and lungs, with their thick teeth adapted for biting through tough bone, as indicated by bite marks inflicted to the rib cage and other tough bony areas on whale remains.[40] Furthermore, attack patterns could differ for prey of different sizes. Fossil remains of some small cetaceans, for example cetotheres, suggest that they were rammed with great force from below before being killed and eaten, based on compression fractures.[68]
During the Pliocene, larger and more advanced cetaceans appeared.[69] Megalodon apparently further refined its hunting strategies to cope with these large whales. Numerous fossilized flipper bones (i.e., segments of the pectoral fins) and caudal vertebrae of large whales from the Pliocene have been found with megalodon bite marks, which suggests that megalodon would immobilize a large whale by ripping apart or biting off its fins before killing and feeding on it.[34][40]
Nursery areas

Megalodon, like contemporaneous sharks, made use of nursery areas to birth their young in, with large amounts of food and protection from predators. Fossil evidence suggests that the preferred nursery sites of megalodon were warm-water coastal environments, where threats were minor and food plentiful.[27] Nursery sites were identified in the Gatún Formation of Panama, the Calvert Formation of Maryland, Banco de Concepción in the Canary Islands,[70] and the Bone Valley Formation of Florida. Given that all extant lamniform sharks give birth to live young, this is believed to have been true of megalodon also.[71] Infant megalodons were around 3.5 meters (11 ft) at their smallest,[23]:61 and the pups were vulnerable to predation by other shark species, such as the great hammerhead shark (Sphyrna mokarran) and the snaggletooth shark (Hemipristis serra).[27] Their dietary preferences display an ontogenetic shift:[23]:65 Young megalodon commonly preyed on fish,[27] giant sea turtles,[55] dugongs,[15]:129 and small cetaceans; mature megalodon moved to off-shore areas and consumed large cetaceans.[23]:74–75
An exceptional case in the fossil record suggests that juvenile megalodon may have occasionally attacked much larger balaenopterid whales. Three tooth marks apparently from a 4-to-7-meter (13 to 23 ft) long Pliocene shark were found on a rib from an ancestral blue or humpback whale that showed evidence of subsequent healing, which is suspected to have been inflicted by a juvenile megalodon.[72][73]
Extinction
Climate change
The Earth experienced a number of changes during the time period megalodon existed which affected marine life. A cooling trend starting in the Oligocene 35 mya ultimately led to glaciation at the poles. Geological events changed currents and precipitation; among these were the closure of the Central American Seaway and changes in the Tethys Ocean, contributing to the cooling of the oceans. The stalling of the Gulf Stream prevented nutrient-rich water from reaching major marine ecosystems, which may have negatively affected its food sources. As its range did not apparently extend into colder waters, megalodon may not have been able to retain a significant amount of metabolic heat, so its range was restricted to shrinking warmer waters.[74][59][75] (This has been disputed; see below.) Fossil evidence confirms the absence of megalodon in regions around the world where water temperatures had significantly declined during the Pliocene.[23]:77 The largest fluctuation of sea levels in the Cenozoic era occurred in the Plio-Pleistocene, between around 5 million to 12 thousand years ago, due to the expansion of glaciers at the pole, which negatively impacted coastal environments, and may have contributed to its extinction along with those of several other marine megafaunal species.[76] These oceanographic changes, in particular the sea level drops, may have restricted many of the suitable shallow warm-water nursery sites for megalodon, hindering reproduction.[74] Nursery areas are pivotal for the survival of many shark species, in part because they protect juveniles from predation.[77][27]
An analysis of the distribution of megalodon over time suggests that temperature change did not play a direct role in its extinction. Its distribution during the Miocene and Pliocene did not correlate with warming and cooling trends; while abundance and distribution declined during the Pliocene, megalodon did show a capacity to inhabit anti-tropical latitudes. It was found in locations with a mean temperature ranging from 12 to 27 °C (54 to 81 °F), with a total range of 1 to 33 °C (34 to 91 °F), indicating that the global extent of suitable habitat should not have been greatly affected by the temperature changes that occurred.[19] This is consistent with evidence that it was a mesotherm.[42]
Changing ecosystem

Marine mammals attained their greatest diversity during the Miocene,[23]:71 such as with baleen whales with over 20 recognized genera in comparison to only six extant genera.[79] Such diversity presented an ideal setting to support a super-predator such as megalodon.[23]:75 By the end of the Miocene, many species of mysticetes had gone extinct;[61] surviving species may have been faster swimmers and thus more elusive prey.[15]:46 Furthermore, after the closure of the Central American Seaway, tropical whales decreased in diversity and abundance.[75] The extinction of megalodon correlates with the decline of many small mysticete lineages, and it is possible that it was quite dependent on them as a food source.[58] Additionally, a marine megafauna extinction during the Pliocene was discovered to have eliminated 36% of all large marine species including 55% of marine mammals, 35% of seabirds, 9% of sharks, and 43% of sea turtles. The extinction was selective for endotherms and mesotherms relative to poikilotherms, implying causation by a decreased food supply[76] and thus consistent with megalodon being mesothermic.[42] Megalodon may have been too large to sustain itself on the declining marine food resources.[74] The cooling of the oceans during the Pliocene might have restricted the access of megalodon to the polar regions, depriving it of access to the large whales which had migrated there.[75]
Competition from new superpredators, such as macropredatory sperm whales which appeared in the Miocene, and killer whales and great white sharks in the Pliocene,[61][65][80] may have also contributed to the decline and extinction of megalodon.[19][15]:46–47[74] Fossil records indicate that the new whale-eating cetaceans commonly occurred at high latitudes during the Pliocene, indicating that they could cope with the increasingly prevalent cold water temperatures; but they also occurred in the tropics (e.g., Orcinus sp. in South Africa).[65]
The extinction of megalodon set the stage for further changes in marine communities. The average body size of baleen whales increased significantly after its disappearance, although possibly due to other, climate-related, causes.[81] Conversely the increase in baleen whale size may have contributed to the extinction of megalodon, as they may have preferred to go after smaller whales; bite marks on large whale species may have come from scavenging sharks. Megalodon may have simply become coextinct with smaller whale species, such as Piscobalaena nana.[78] The extinction of megalodon had a positive impact on other apex predators of the time, such as the great white shark, in some cases spreading to regions where megalodon became absent.[19][80][82]
In popular culture

Megalodon has been portrayed in several works of fiction, including films and novels, and continues to be a popular subject for fiction involving sea monsters.[83] Three individual megalodon, two adults and one juvenile, were portrayed in BBC's 2003 TV series Sea Monsters, where it is defined as a "hazard" of the era.[84] The History Channel's Jurassic Fight Club portrays a megalodon attacking a Brygmophyseter sperm whale in Japan.[85] Several films depict megalodon, such as Shark Attack 3: Megalodon and the Mega Shark series (for instance Mega Shark Versus Giant Octopus and Mega Shark Versus Crocosaurus).[83] The shark appears in the 2017 videogame Ark: Survival Evolved.[86] Some stories, such as Jim Shepard's Tedford and the Megalodon, portray a rediscovery of the shark.[87] Steve Alten's Meg: A Novel of Deep Terror portrays the shark having preyed on dinosaurs with its prologue and cover artwork depicting megalodon killing a Tyrannosaurus in the sea.[88] The sequels to the book also star megalodon: The Trench, Meg: Primal Waters, Meg: Hell's Aquarium, and Meg: Origins,[83] and there is a film adaptation entitled The Meg released on August 10, 2018.[89]
Animal Planet's pseudo-documentary Mermaids: The Body Found included an encounter 1.6 mya between a pod of mermaids and a megalodon.[90] Later, in August 2013, the Discovery Channel opened its annual Shark Week series with another film for television, Megalodon: The Monster Shark Lives,[91] a controversial docufiction about the creature that presented alleged evidence in order to suggest that megalodon was still alive. This program received criticism for being completely fictional; for example, all of the supposed scientists depicted were paid actors. In 2014, Discovery re-aired The Monster Shark Lives, along with a new one-hour program, Megalodon: The New Evidence, and an additional fictionalized program entitled Shark of Darkness: Wrath of Submarine, resulting in further backlash from media sources and the scientific community.[40][92][93]
Reports of supposedly fresh megalodon teeth, such as those made by HMS Challenger in 1873 which were erroneously dated to be around 11,000 to 24,000 years old, are probably teeth that were well-preserved by a thick mineral-crust precipitate of manganese dioxide, and so had a lower decomposition rate and retained a white color during fossilization. Fossil megalodon teeth can vary in color from off-white to dark browns and greys, and some fossil teeth may have been redeposited into a younger stratum. The claims that megalodon could remain elusive in the depths, similar to the megamouth shark which was discovered in 1976, are unlikely as the shark lived in warm coastal waters and probably could not survive in the cold and nutrient-poor deep sea environment.[94]
See also
References
- 1 2 Agassiz, Louis (1843). Recherches sur les poissons fossiles [Research on the fossil fishes] (in French). Neuchatel: Petitpierre. p. 41.
- ↑ "Otodus (Megaselachus) megalodon (Agassiz, 1837)". SharkReferences.com. Retrieved 24 October 2017.
- ↑ Eastman, C. R. (1904). Maryland Geological Survey. 2. Baltimore, Maryland: Johns Hopkins University. p. 82.
- 1 2 3 Cappetta, H. (1987). "Mesozoic and Cenozoic Elasmobranchii". Handbook of Paleoichthyology. 3B. München, Germany: Friedrich Pfeil. ISBN 978-3-89937-046-1. OCLC 829906016.
- ↑ Hay, O. P. (1901). "Bibliography and Catalogue of the Fossil Vertebrata of North America". Bulletin of the United States Geological Society (179): 308.
- ↑ https://www.britannica.com/animal/megalodon
- ↑ Haven, Kendall (1997). 100 Greatest Science Discoveries of All Time. Westport, Connecticut: Libraries Unlimited. pp. 25–26. ISBN 978-1-59158-265-6. OCLC 230807846.
- ↑ Kuang-Tai, Hsu (2009). "The Path to Steno's Synthesis on the Animal Origin of Glossopetrae". In Rosenburg, G. D. The Revolution in Geology from the Renaissance to the Enlightenment. 203. Boulder, Colorado: Geological Society of America. ISBN 978-0-8137-1203-1. OCLC 608657795.
- ↑ Eilperin, J. (2012). Demon Fish. Pantheon Books. p. 43. ISBN 978-0-7156-4352-5.
- 1 2 3 4 5 6 Nyberg, K. G.; Ciampaglio C. N.; Wray G. A. (2006). "Tracing the ancestry of the great white shark, Carcharodon carcharias, using morphometric analyses of fossil teeth". Journal of Vertebrate Paleontology. 26 (4): 806–814. doi:10.1671/0272-4634(2006)26[806:TTAOTG]2.0.CO;2.
- ↑ Keyes, I. W. (2012). "New records of the Elasmobranch C. megalodon (Agassiz) and a review of the genus Carcharodon in the New Zealand fossil record". New Zealand Journal of Geology and Geophysics. 15 (2): 229. doi:10.1080/00288306.1972.10421956.
- ↑ μέγας. Liddell, Henry George; Scott, Robert; A Greek–English Lexicon at the Perseus Project
- ↑ ὀδούς. Liddell, Henry George; Scott, Robert; A Greek–English Lexicon at the Perseus Project
- 1 2 Augilera, Orangel A.; García, Luis; Cozzuol, Mario A. (2008). "Giant-toothed white sharks and cetacean trophic interaction from the Pliocene Caribbean Paraguaná Formation". Paläontologische Zeitschrift. 82 (2): 204–208. doi:10.1007/BF02988410. ISSN 0038-2353.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 Renz, Mark (2002). Megalodon: Hunting the Hunter. Lehigh Acres, Florida: PaleoPress. pp. 1–159. ISBN 978-0-9719477-0-2. OCLC 52125833.
- 1 2 3 4 Ehret D. J.; Hubbell G.; Macfadden B. J. (2009). "Exceptional preservation of the white shark Carcharodon from the early Pliocene of Peru". Journal of Vertebrate Paleontology. 29 (1): 1–13. doi:10.1671/039.029.0113. JSTOR 20491064.
- ↑ Hideo, Habe; Mastatoshi, Goto; Naotomo, Kaneko (2004). "Age of Carcharocles megalodon (Lamniformes: Otodontidae): A review of the stratigraphic records". The Palaeontological Society of Japan. 75 (75): 7–15.
- 1 2 3 4 Gottfried, M. D.; Fordyce, R. E. (2001). "An associated specimen of Carcharodon angustidens (Chondrichthyes, Lamnidae) from the Late Oligocene of New Zealand, with comments on Carcharodon interrelationships". Journal of Vertebrate Paleontology. 21 (4): 730–739. doi:10.1671/0272-4634(2001)021[0730:AASOCA]2.0.CO;2.
- 1 2 3 4 5 6 7 8 9 10 11 Pimiento, C.; MacFadden, B. J.; Clements, C. F.; Varela, S.; Jaramillo, C.; Velez-Juarbe, J.; Silliman, B. R. (2016). "Geographical distribution patterns of Carcharocles megalodon over time reveal clues about extinction mechanisms". Journal of Biogeography. 43 (8): 1645–1655. doi:10.1111/jbi.12754.
- 1 2 3 4 Pimiento, C.; Clements, C. F. (2014). "When Did Carcharocles megalodon Become Extinct? A New Analysis of the Fossil Record". PLoS ONE. 9 (10): e111086. Bibcode:2014PLoSO...9k1086P. doi:10.1371/journal.pone.0111086. PMC 4206505. PMID 25338197.
- 1 2 3 Shimada, K.; Chandler, R. E.; Lam, O. L. T.; Tanaka, T.; Ward, D. J. (2016). "A new elusive otodontid shark (Lamniformes: Otodontidae) from the lower Miocene, and comments on the taxonomy of otodontid genera, including the 'megatoothed' clade". Historical Biology. 29 (5): 1–11. doi:10.1080/08912963.2016.1236795.
- 1 2 3 4 Pimiento, C.; Balk, M. A. (2015). "Body-size trends of the extinct giant shark Carcharocles megalodon: a deep-time perspective on marine apex predators". Paleobiology. 41 (3): 479–490. doi:10.1017/pab.2015.16. PMC 4541548. PMID 26321775.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 Klimley, Peter; Ainley, David (1996). "Evolution". Great White Sharks: The Biology of Carcharodon carcharias. San Diego, California: Academic Press. ISBN 978-0-12-415031-7. OCLC 212425118.
- 1 2 3 4 5 Andres, Lutz (2002). "C. megalodon — Megatooth Shark, Carcharodon versus Carcharocles". fossilguy.com. Retrieved 16 January 2008.
- ↑ Siverson, Mikael; Johan Lindgren; Michael G. Newbrey; Peter Cederström; Todd D. Cook (2013). "Late Cretaceous (Cenomanian-Campanian) mid-palaeolatitude sharks of Cretalamna appendiculata type" (PDF). Acta Palaeontologica Polonica: 2. doi:10.4202/app.2012.0137. Archived from the original on 19 October 2013.
- ↑ Benton, M. J.; Pearson, P. N. (2001). "Speciation in the fossil record". Trends in Ecology and Evolution. 16 (7): 405–411. doi:10.1016/s0169-5347(01)02149-8. PMID 11403874.
- 1 2 3 4 5 6 7 8 9 Pimiento, Catalina; Ehret, Dana J.; MacFadden, Bruce J.; Hubbell, Gordon (2010). Stepanova, Anna, ed. "Ancient Nursery Area for the Extinct Giant Shark Megalodon from the Miocene of Panama". PLoS ONE. 5 (5): e10552. Bibcode:2010PLoSO...510552P. doi:10.1371/journal.pone.0010552. PMC 2866656. PMID 20479893.
- ↑ Lawley, R. (1881). "Selache manzonii n. sp. – Dente Fossile délia Molassa Miocenica del Monte Titano (Repubblica di San Marino)" [Fossil tooth from Miocene Molasse from Monte Titano (Republic of San Marino)]. Atti della Società Toscana di Scienze Naturali (in Italian). 5: 167–172.
- ↑ "Could Megalodon Have Looked Like a BIG Sandtiger Shark?". Biology of Sharks and Rays. Retrieved 2 September 2017.
- ↑ Portell, Roger; Hubell, Gordon; Donovan, Stephen; Green, Jeremy; Harper, David; Pickerill, Ron (2008). "Miocene sharks in the Kendeace and Grand Bay formations of Carriacou, The Grenadines, Lesser Antilles" (PDF). Caribbean Journal of Science. 44 (3): 279–286. doi:10.18475/cjos.v44i3.a2. Archived from the original on 20 July 2011.
- 1 2 Randall, John (July 1973). "Size of the Great White Shark (Carcharodon)". Science Magazine. 181 (4095): 169–170. Bibcode:1973Sci...181..169R. doi:10.1126/science.181.4095.169. PMID 17746627.
- ↑ Schembri, Patrick (1994). "Malta's Natural Heritage". Natural Heritage. In: 105–124.
- ↑ Papson, Stephen (1992). "Copyright: Cross the Fin Line of Terror". Journal of American Culture. 15 (4): 67–81. doi:10.1111/j.1542-734X.1992.1504_67.x.
- 1 2 3 4 5 Wroe, S.; Huber, D. R.; Lowry, M.; McHenry, C.; Moreno, K.; Clausen, P.; Ferrara, T. L.; Cunningham, E.; Dean, M. N.; Summers, A. P. (2008). "Three-dimensional computer analysis of white shark jaw mechanics: how hard can a great white bite?" (PDF). Journal of Zoology. 276 (4): 336–342. doi:10.1111/j.1469-7998.2008.00494.x.
- ↑ Compagno, Leonard J. V. (2002). Sharks of the World: An Annotated and Illustrated Catalogue of Shark Species Known to Date. Rome: Food & Agriculture Organization of the United Nations. p. 97. ISBN 978-92-5-104543-5.
- ↑ "Great white sharks: 10 myths debunked". The Guardian. Retrieved 3 June 2016.
- ↑ Carpenter, K. "Carcharodon carcharias". FishBase.org. Retrieved 3 June 2016.
- ↑ Viegas, Jennifer. "Largest Great White Shark Don't Outweigh Whales, but They Hold Their Own". Discovery Channel. Retrieved 19 January 2010.
- ↑ Wood, Gerald L. (1976). The Guinness Book of Animal Facts and Feats. Guinness Superlatives. pp. 139–141. ISBN 978-0-900424-60-1.
- 1 2 3 4 5 6 7 8 Prothero, D. R. (2015). "Mega-Jaws". The Story of Life in 25 Fossils. New York, New York: Columbia University Press. pp. 96–110. ISBN 978-0-231-17190-8. OCLC 897505111.
- ↑ Jacoby, D. M. P.; Siriwat, P.; Freeman, R.; Carbone, C. (2015). "Is the scaling of swim speed in sharks driven by metabolism?". Biology Letters. 12 (10): 20150781. doi:10.1098/rsbl.2015.0781. PMC 4707698. PMID 26631246.
- 1 2 3 Ferrón, H. G. (2017). "Regional endothermy as a trigger for gigantism in some extinct macropredatory sharks". PLOS ONE. 12 (9): e0185185. Bibcode:2017PLoSO..1285185F. doi:10.1371/journal.pone.0185185. PMC 5609766. PMID 28938002.
- ↑ Crane, B. (2017). "A Prehistoric Killer, Buried in Muck". The New Yorker. Retrieved 10 December 2017.
- ↑ Mustain, A. (2011). "For Sale: World's Largest Shark Jaws". LiveScience. Retrieved 31 August 2017.
- ↑ Helfman, G.; Burgess, G. H. (2014). Sharks: The Animal Answer Guide. Baltimore, Maryland: Johns Hopkins University Press. p. 19. ISBN 978-1-4214-1310-5. OCLC 903293986.
- ↑ Shimada, Kenshu (2002). "The relationship between the tooth size and total body length in the white shark, Carcharodon carcharias (Lamniformes: Lamnidae)". Journal of Fossil Research. 35 (2): 28–33.
- 1 2 Pimiento, Catalina; Gerardo González-Barba; Dana J. Ehret; Austin J. W. Hendy; Bruce J. MacFadden; Carlos Jaramillo (2013). "Sharks and Rays (Chondrichthyes, Elasmobranchii) from the Late Miocene Gatun Formation of Panama" (PDF). Journal of Paleontology. 87 (5): 755–774. doi:10.1666/12-117. Archived from the original (PDF) on 29 October 2013.
- 1 2 Bendix-Almgreen, Svend Erik (1983). "Carcharodon megalodon from the Upper Miocene of Denmark, with comments on elasmobranch tooth enameloid: coronoïn" (PDF). Bulletin of the Geological Society of Denmark. 32: 1–32.
- ↑ Reolid, M.; Molina, J. M. (2015). "Record of Carcharocles megalodon in the Eastern Guadalquivir Basin (Upper Miocene, South Spain)". Estudios Geológicos. 71 (2): e032. doi:10.3989/egeol.41828.342.
- ↑ Uyeno, T.; Sakamoto, O.; Sekine, H. (1989). "The Description of an Almost Complete Tooth Set of Carcharodon megalodon from a Middle Miocene Bed in the Saitama Prefecture, Japan". Saitama Museum of Natural History Bulletin. 7: 73–85.
- ↑ Anderson, P.S.L.; Westneat, M. (2009). "A biomechanical model of feeding kinematics for Dunkleosteus terrelli (Arthrodira, Placodermi)". Paleobiology. 35 (2): 251–269. doi:10.1666/08011.1.
- ↑ "Megalodon Shark Facts and Information: The Details". fossilguy.com. Retrieved 18 September 2017.
- ↑ Stringer, G. L.; King, L. (2012). "Late Eocene Shark Coprolites from the Yazoo Clay in Northeastern Louisiana". New Mexico Museum of Natural History and Science, Bulletin 57. Vertebrate Corpolites: 301.
- ↑ Fitzgerald, Erich (2004). "A review of the Tertiary fossil Cetacea (Mammalia) localities in Australia" (PDF). Memoirs of Museum Victoria. 61 (2): 183–208. Archived from the original (PDF) on 23 August 2008.
- 1 2 3 Aguilera O.; Augilera E. R. D. (2004). "Giant-toothed White Sharks and Wide-toothed Mako (Lamnidae) from the Venezuela Neogene: Their Role in the Caribbean, Shallow-water Fish Assemblage". Caribbean Journal of Science. 40 (3): 362–368.
- 1 2 Carcharocles megalodon at fossilworks.org (retrieved 28 August 2017)
- ↑ Martin, J. E.; Tacail, T.; Sylvain, A.; Catherine, G.; Vincent, B. (2015). "Calcium isotopes reveal the trophic position of extant and fossil elasmobranchs". Chemical Geology. 415: 118–125. Bibcode:2015ChGeo.415..118M. doi:10.1016/j.chemgeo.2015.09.011.
- 1 2 3 Collareta, A.; Lambert, O.; Landini, W.; Di Celma, C.; Malinverno, E.; Varas-Malca, R.; Urbina, M.; Bianucci, G. (2017). "Did the giant extinct shark Carcharocles megalodon target small prey? Bite marks on marine mammal remains from the late Miocene of Peru". Palaeogeography, Palaeoclimatology, Palaeoecology. 469: 84–91. doi:10.1016/j.palaeo.2017.01.001.
- 1 2 Morgan, Gary S. (1994). "Whither the giant white shark?" (PDF). Paleontology Topics. 2 (3): 1–2. Archived from the original (PDF) on 22 July 2016.
- ↑ Landini, W.; Altamirano-Sera, A.; Collareta, A.; Di Celma, C.; Urbina, M.; Bianucci, G. (2017). "The late Miocene elasmobranch assemblage from Cerro Colorado (Pisco Formation, Peru)". Journal of South American Earth Sciences. 73: 168–190. Bibcode:2017JSAES..73..168L. doi:10.1016/j.jsames.2016.12.010.
- 1 2 3 4 5 6 7 8 Lambert, O.; Bianucci, G.; Post, P.; de Muizon, C.; Salas-Gismondi, R.; Urbina, M.; Reumer, J. (2010). "The giant bite of a new raptorial sperm whale from the Miocene epoch of Peru". Nature. 466 (7302): 105–108. Bibcode:2010Natur.466..105L. doi:10.1038/nature09067. PMID 20596020.
- ↑ Compagno, Leonard J. V. (1989). "Alternative life-history styles of cartilaginous fishes in time and space". Environmental Biology of Fishes. 28 (1–4): 33–75. doi:10.1007/BF00751027.
- ↑ Ferretti, Francesco; Boris Worm; Gregory L. Britten; Michael R. Heithaus; Heike K. Lotze1 (2010). "Patterns and ecosystem consequences of shark declines in the ocean" (PDF). Ecology Letters. 13 (8): 1055–1071. doi:10.1111/j.1461-0248.2010.01489.x. PMID 20528897. Archived from the original on 6 July 2011. Retrieved 19 February 2011.
- ↑ Bianucci, Giovanni; Walter, Landini (2006). "Killer sperm whale: a new basal physeteroid (Mammalia, Cetacea) from the Late Miocene of Italy". Zoological Journal of the Linnean Society. 148 (1): 103–131. doi:10.1111/j.1096-3642.2006.00228.x.
- 1 2 3 Heyning, John; Dahlheim, Marilyn (1988). "Orcinus orca" (PDF). Mammalian Species. 304 (304): 1–9. doi:10.2307/3504225. JSTOR 3504225. Archived from the original (PDF) on 5 December 2010.
- ↑ "Paleoecology of Megalodon and the White Shark". Biology of Sharks and Rays. Retrieved 1 October 2017.
- ↑ Tanke, Darren; Currie, Philip (1998). "Head-Biting Behaviour in Theropod Dinosaurs: Paleopathological Evidence" (PDF). Gaia N°15: 168.
- 1 2 Godfrey, S. J.; Altman, J. (2005). "A Miocene Cetacean Vertebra Showing a Partially Healed Compression Factor, the Result of Convulsions or Failed Predation by the Giant White Shark, Carcharodon megalodon" (PDF). Jeffersoniana (16): 1–12.
- ↑ Deméré, Thomas A.; Berta, Annalisa; McGowen, Michael R. (2005). "The taxonomic and evolutionary history of fossil and modern balaenopteroid mysticetes". Journal of Mammalian Evolution. 12 (1/2): 99–143. doi:10.1007/s10914-005-6944-3.
- ↑ "Identifican en Canarias fósiles de 'megalodón', el tiburón más grande que ha existido" [Identifying Canary fossils of 'megalodon', the largest shark that ever lived] (in Spanish). Europa Press Noticias SA. 2013. Retrieved 29 August 2017.
- ↑ Dulvy, N. K.; Reynolds, J. D. (1997). "Evolutionary transitions among egg-laying, live-bearing and maternal inputs in sharks and rays". Proceedings of the Royal Society B: Biological Sciences. 264 (1386): 1309–1315. doi:10.1098/rspb.1997.0181. PMC 1688595.
- ↑ Godfrey, Stephen (2004). "The Ecphora" (PDF). The Newsletter of Calvert Marine Museum Fossil Club. 19 (1): 1–13. Archived from the original on 10 December 2010.
- ↑ Kallal, R. J.; Godfrey, S. J.; Ortner, D. J. (27 August 2010). "Bone Reactions on a Pliocene Cetacean Rib Indicate Short-Term Survival of Predation Event". International Journal of Osteoarchaeology. 22 (3): 253–260. doi:10.1002/oa.1199.
- 1 2 3 4 "The Extinction of Megalodon". Biology of Sharks and Rays. Retrieved 31 August 2017.
- 1 2 3 Allmon, Warren D.; Steven D. Emslie; Douglas S. Jones; Gary S. Morgan (2006). "Late Neogene Oceanographic Change along Florida's West Coast: Evidence and Mechanisms". The Journal of Geology. 104 (2): 143–162. Bibcode:1996JG....104..143A. doi:10.1086/629811.
- 1 2 Pimiento, C.; Griffin, J. N.; Clements, C. F.; Silvestro, D.; Varela, S.; Uhen, M. D.; Jaramillo, C. (2017). "The Pleistocene Marine Megafauna Extinction and its Impact on Functional Diversity". Nature Ecology and Evolution. 1 (8): 1100–1106. doi:10.1038/s41559-017-0223-6. PMID 29046566.
- ↑ Reilly, Michael (29 September 2009). "Prehistoric Shark Nursery Spawned Giants". Discovery News. Archived from the original on 10 March 2012. Retrieved 23 November 2013.
- 1 2 Collareta, A.; Lambert, O.; Landini, W.; Bianucci, G. (2017). "Did the giant extinct shark Carcharocles megalodon target small prey? Bite marks on marine mammal remains from the late Miocene of Peru". Palaeogeography, Palaeoclimatology, Palaeoecology. 469: 84–91. doi:10.1016/j.palaeo.2017.01.001.
- ↑ Dooly A.C.; Nicholas C. F.; Luo Z. X. (2006). "The earliest known member of the rorqual—gray whale clade (Mammalia, Cetacea)". Journal of Vertebrate Paleontology. 24 (2): 453–463. doi:10.1671/2401. JSTOR 4524731.
- 1 2 Antunes, Miguel Telles; Balbino, Ausenda Cáceres (2010). "The Great White Shark Carcharodon carcharias (Linne, 1758) in the Pliocene of Portugal and its Early Distribution in Eastern Atlantic". Revista Española de Paleontología. 25 (1): 1–6.
- ↑ Slater, G. J.; Goldbogen, J. A.; Pyenson, N. D. (2017). "Independent evolution of baleen whale gigantism linked to Plio-Pleistocene ocean dynamics". Proceedings of the Royal Society B: Biological Sciences. 284 (1855): 20170546. doi:10.1098/rspb.2017.0546. PMC 5454272. PMID 28539520.
- ↑ Sylvain, Adnet; A. C. Balbino; M. T. Antunes; J. M. Marín-Ferrer (2010). "New fossil teeth of the White Shark (Carcharodon carcharias) from the Early Pliocene of Spain. Implication for its paleoecology in the Mediterranean". Neues Jahrbuch für Geologie und Paläontologie. 256 (1): 7–16. doi:10.1127/0077-7749/2009/0029.
- 1 2 3 Weinstock, J. A. (2014). The Ashgate Encyclopedia of Literary and Cinematic Monsters. Farnham, United Kingdom: Routledge. pp. 107–108. ISBN 978-1-4094-2562-5. OCLC 874390267.
- ↑ "The Third Most Deadly Sea". Sea Monsters. Season 1. Episode 3. 23 September 2003. BBC.
- ↑ "Deep Sea Killers". Jurassic Fight Club. Season 1. Episode 5. 26 August 2008. History Channel.
- ↑ "Megalodon". ARK Survival Evolved. Retrieved 9 August 2018.
- ↑ Shepard, J. (2007). "Tedford and the Megalodon". In Chabon, M. McSweeney's Mammoth Treasury of Thrilling Tales. New York, New York: Knopf Doubleday Publishing Group. p. 9. ISBN 978-0-307-42682-6. OCLC 1002088939.
- ↑ Alten, S. (2011). "Megalodon". Meg: A Novel of Deep Terror. Portland, Oregon: Gere Donovan Press. ISBN 978-1-936666-21-8.
- ↑ McNary, Dave (2 March 2017). "Jason Statham's Shark Thriller 'Meg' Swims Back Five Months". Variety. Retrieved 15 April 2018.
- ↑ Sid Bennett (director) (27 May 2012). Mermaids: The Body Found (Motion picture). Animal Planet.
- ↑ "Shark Week 'Megalodon: The Monster Shark Lives' Tries To Prove Existence Of Prehistoric Shark (VIDEO)". Huff Post Green. 5 August 2013. Retrieved 11 August 2013.
- ↑ Winston, B.; Vanstone, G.; Chi, W. (2017). "A Walk in the Woods". The Act of Documenting: Documentary Film in the 21st Century. New York, New York: Bloomsbury Publishing. ISBN 978-1-5013-0918-2. OCLC 961183719.
- ↑ Flanagin, J. (2014). "Sorry, Fans. Discovery Has Jumped the Shark Week". New York Times. Retrieved 16 August 2014.
- ↑ "Does Megalodon Still Live?". Biology of Sharks and Rays. Retrieved 2 October 2017.
Further reading
- Dickson, K. A.; Graham, J. B. (November–December 2004). "Evolution and consequences of endothermy in fishes". Physiological and Biochemical Zoology. 77 (6): 998–1018. doi:10.1086/423743. PMID 15674772.
- Kent, Bretton W. (1994). Fossil Sharks of the Chesapeake Bay Region. Columbia, Md.: Egan Rees & Boyer. ISBN 978-1-881620-01-3. OCLC 918266672.
External links
| Wikimedia Commons has media related to Carcharodon megalodon. |
| Wikispecies has information related to Megalodon |
- Paleontological videos
Note: Flash Player is required to view the content below.
- Paleontologist Mark Renz shows one of the largest megalodon teeth discovered on YouTube
- Shark Week Special on megalodon with Pat McCarthy and John Babiarz on YouTube with comments on its extinction.
- Megalodon fossil teeth show evidence of 10-million-year-old shark nursery on YouTube
- Expert view: information about megalodon on YouTube (featuring expert Dana Ehret)
- Lamniform sharks: 110 million years of ocean supremacy on YouTube (featuring expert Mikael Siverson)
- The Rise and Fall of the Neogene Giant Sharks on YouTube (featuring expert Bretton Kent)
| Topics | |
|---|---|
| Taxonomy |
|
| Human interaction | |
| |