DNA demethylation
DNA demethylation is the process of removal of a methyl group from nucleotides in DNA. Both DNA demethylation and methylation play important roles in mammalian development and differentiation, as well as in cognition and neuroregeneration (after injury to peripheral nerves in mammals). DNA methylation on cytosine at CpG sites on a gene promoter leads to the silencing of gene expression, while DNA demethylation of a gene promoter is linked to transcriptional activation and gene expression. These are called epigenetic changes. The DNA methylation status that controls gene expression is copied during DNA replication and is transmitted to daughter cells along with the DNA sequence. This area is currently being investigated for its role in disease progression and for potential treatments, such as cancer therapy.[1]
Passive and active demethylation
DNA demethylation can occur through passive or active mechanisms. The passive process takes place in the absence of methylation of newly synthesised DNA strands by DNMT1 during several replication rounds (for example, upon 5-Azacytidine treatment), leading to dilution of the methylation signal. Active DNA demethylation is mediated by multiple enzymes and can occur independent of DNA replication.
Examples DNA Demethylation
All the cases of DNA demethylation can be classified as global (genome wide) or locus-specific (when just specific sequences are demethylated). The genome-wide DNA demethylation occurs:
- In mammals:
- In the male pronucleus of zygote immediately after fertilization;
- In mouse primordial germ cells (PGCs) between E8.5-11.5 day old embryos;[2]
- Possibly in amphibia - during midblastula transition.
Examples of specific DNA demethylation:
- Genomic imprinting during plant reproduction;
- Electroconvulsive stimulation-induced demethylation of neurotrophic factor genes in dentate gyrus neurons in the mouse brain.[3][4]
Possible mechanisms of active DNA demethylation
There are several proposed hypothetical mechanisms of active DNA demethylation:
A Direct removal of 5-methylcytosine
- Direct removal of methyl group. This process has quite low thermodynamic probability.
- Removal of methylated bases (either by direct removal of methylcytosine, or through cytosine deamination followed by removal of thymine from thymine/guanosine mismatch), followed by insertion of unmethylated one using base excision repair machinery (BER).
- Removal of entire DNA patch and following filling it with new nucleotides by nucleotide excision repair (NER) or mismatch repair (MMR).[5]
B Removal of 5-methylcytosine via further modified cytosine bases
Oxidation of the methyl group generates 5-Hydroxymethylcytosine. Several mechanisms have been proposed to mediate demethylation of 5-hydroxymethylcytosines.[6][7] This base can be either deaminated by AID/Apobec enzymes to give 5-Hydroxymethyluracil.[4] Alternatively, TET enzymes can further oxidize 5-hydroxymethylcytosine to 5-Formylcytosine and 5-Carboxylcytosine.[8][9][10]
- Both the deamination and the oxidation products have been shown to be repaired by TDG, a glycosylase which is involved in base excision repair.[9][11][12] A base excision mediated demethylation mechanism would yield double strand breaks if it occurs on large scale in CpG islands.
- The carboxyl and formyl groups of 5-Formylcytosine and 5-Carboxylcytosine could be enzymatically removed without excision of the base.[6][7][8][10] Precedent for similar reactions is found in biosynthetic pathways.
DNA hydroxymethylation
DNA hydroxymethylation has been proposed to act as a specific epigenetic mark opposing DNA methylation, rather than a passive intermediate in the de-methylation pathway. DNA hydroxymethylation in vivo is sometimes associated with labile nucleosomes, which are more easy to disassemble and to be out-competed by transcription factors during cell development.[13] Hydroxymethylation has been associated with pluripotency of stem cells. Furthermore, changes in hydroxymethylation have been associated with cancer.[14]
Cognition
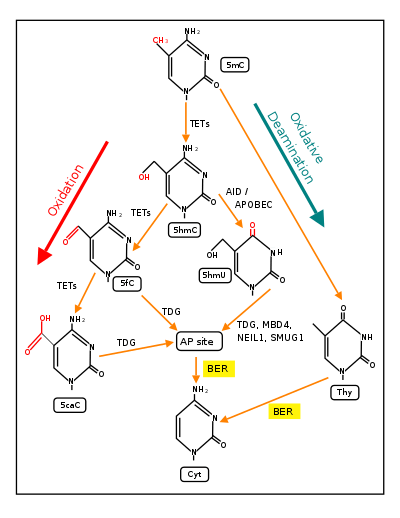
Active DNA methylation and demethylation is required for the cognition process of memory formation and maintenance.[16] In rats, contextual fear conditioning can trigger life-long memory for the event with a single trial, and methylation changes appear to be correlated with triggering particularly long-lived memories.[16] With contextual fear conditioning, after 24 hours, DNA isolated from the rat brain hippocampus region had 2097 differentially methylated genes, with a portion being demethylated.[16] As reviewed by Bayraktar and Kreutz,[15] in the brain, DNA demethylation is dependent upon the steps indicated in the figure and depends on base excision repair (BER) as the terminal step (see figure).
Physical exercise has well established beneficial effects on learning and memory (see Neurobiological effects of physical exercise). BDNF is a particularly important regulator of learning and memory.[17] As reviewed by Fernandes et al.,[18] in rats, exercise enhances the hippocampus expression of the gene Bdnf, which has an essential role in memory formation. Enhanced expression of Bdnf occurs through demethylation of its CpG island promoter at exon IV[18] and this demethylation depends on steps illustrated in the figure.[15]
Peripheral sensory neuron regeneration
After injury, neurons in the adult peripheral nervous system can switch from a dormant state with little axonal growth to robust axon regeneration. DNA demethylation in mature mammalian neurons removes barriers to axonal regeneration.[19] This demethylation, in regenerating mouse peripheral neurons, depends upon TET3 to generate 5-hydroxymethylcytosine (5hmC) in DNA.[19][20] 5hmC was altered in a large set of regeneration-associated genes (RAGs), including well-known RAGs such as Atf3, Bdnf, and Smad1, that regulate the axon growth potential of neurons.[20]
See also
References
- ↑ "DNA demethylation". Epigentek. Retrieved 1 August 2018.
- ↑ Hackett, JA; Sengupta, R; Zylicz, JJ; Murakami, K; Lee, C; Down, T; Surani, MA (2012-12-06). "Germline DNA demethylation dynamics and imprint erasure through 5-hydroxymethylcytosine". Science. 339 (6118): 448–52. doi:10.1126/science.1229277. PMC 3847602. PMID 23223451.
- ↑ Ma, DK; Jang, MH; Guo, JU; Kitabatake, Y; Chang, ML; Pow-Anpongkul, N; Flavell, RA; Lu, B; Ming, GL; Song, H (2009-02-20). "Neuronal activity-induced Gadd45b promotes epigenetic DNA demethylation and adult neurogenesis". Science. 323 (5917): 1074–7. doi:10.1126/science.1166859. PMC 2726986. PMID 19119186.
- 1 2 Guo, JU; Su, Y; Zhong, C; Ming, GL; Song, H (2011-04-29). "Hydroxylation of 5-Methylcytosine by TET1 Promotes Active DNA Demethylation in the Adult Brain". Cell. 145 (3): 423–34. doi:10.1016/j.cell.2011.03.022. PMC 3088758. PMID 21496894.
- ↑ Grin, I; Ishchenko, AA (May 2016). "An interplay of the base excision repair and mismatch repair pathways in active DNA demethylation". Nucleic Acids Res. 44 (8): 3713–27. doi:10.1093/nar/gkw059. PMC 4856981. PMID 26843430.
- 1 2 Wu, SC; Zhang, Y (Sep 2010). "Active DNA demethylation: many roads lead to Rome". Nature Reviews Molecular Cell Biology. 11 (9): 607–20. doi:10.1038/nrm2950. PMC 3711520. PMID 20683471.
- 1 2 Globisch, Daniel; Münzel, Martin; Müller, Markus; Michalakis, Stylianos; Wagner, Mirko; Koch, Susanne; Brückl, Tobias; Biel, Martin; Carell, Thomas (23 December 2010). Croft, Anna Kristina, ed. "Tissue Distribution of 5-Hydroxymethylcytosine and Search for Active Demethylation Intermediates". PLoS ONE. 5 (12): e15367. doi:10.1371/journal.pone.0015367. PMC 3009720. PMID 21203455.
- 1 2 Pfaffeneder, Toni; Hackner, Benjamin; Truss, Matthias; Münzel, Martin; Müller, Markus; Deiml, Christian A.; Hagemeier, Christian; Carell, Thomas (30 June 2011). "The Discovery of 5-Formylcytosine in Embryonic Stem Cell DNA". Angew. Chem. Int. Ed. 50 (31): 7008–7012. doi:10.1002/anie.201103899. PMID 21721093.
- 1 2 He, YF; Li, BZ; Li, Z; Liu, P; Wang, Y; Tang, Q; Ding, J; Jia, Y; Chen, Z; Li, L; Sun, Y; Li X; Dai, Q; Song, CX; Zhang, K; He, C; Xu, GL (4 August 2011). "Tet-Mediated Formation of 5-Carboxylcytosine and Its Excision by TDG in Mammalian DNA". Science. 333 (6047): 1303–1307. doi:10.1126/science.1210944. PMC 3462231. PMID 21817016.
- 1 2 Ito, S; Li, S; Dai, Q; Wu, SC; Collins, SB; Swenberg, JA; He, C; Zhang, Y (21 July 2011). "Tet Proteins Can Convert 5-Methylcytosine to 5-Formylcytosine and 5-Carboxylcytosine". Science. 333 (6047): 1300–1303. doi:10.1126/science.1210597. PMC 3495246. PMID 21778364.
- ↑ Maiti, A; Drohat, AC (23 August 2011). "Thymine DNA Glycosylase Can Rapidly Excise 5-Formylcytosine and 5-Carboxylcytosine". J. Biol. Chem. 286 (41): 35334–8. doi:10.1074/jbc.C111.284620. PMC 3195571. PMID 21862836.
- ↑ Cannon, SV; Cummings, GW; Teebor, GW (1988). "5-Hydroxymethylcytosine DNA Glycosylase Activity in Mammalian Tissue". Biochem. Biophys. Res. Commun. 151 (3): 1173–1179. doi:10.1016/S0006-291X(88)80489-3. PMID 3355548.
- ↑ Teif, Vladimir; Beshnova, Daria A.; Vainshtein, Yevhen; Marth, Caroline; Mallm, Jan-Philipp; Höfer, Thomas; Rippe, Karsten (8 May 2014). "Nucleosome repositioning links DNA (de)methylation and differential CTCF binding during stem cell development". Genome Research. 24 (8): 1285–1295. doi:10.1101/gr.164418.113. PMC 4120082. PMID 24812327.
- ↑ http://www.ks.uiuc.edu/Research/methylation/
- 1 2 3 Bayraktar G, Kreutz MR (2018). "The Role of Activity-Dependent DNA Demethylation in the Adult Brain and in Neurological Disorders". Front Mol Neurosci. 11: 169. doi:10.3389/fnmol.2018.00169. PMC 5975432. PMID 29875631.
- 1 2 3 Duke CG, Kennedy AJ, Gavin CF, Day JJ, Sweatt JD (July 2017). "Experience-dependent epigenomic reorganization in the hippocampus". Learn. Mem. 24 (7): 278–288. doi:10.1101/lm.045112.117. PMC 5473107. PMID 28620075.
- ↑ Karpova NN (January 2014). "Role of BDNF epigenetics in activity-dependent neuronal plasticity". Neuropharmacology. 76 Pt C: 709–18. doi:10.1016/j.neuropharm.2013.04.002. PMID 23587647.
- 1 2 Fernandes J, Arida RM, Gomez-Pinilla F (September 2017). "Physical exercise as an epigenetic modulator of brain plasticity and cognition". Neurosci Biobehav Rev. 80: 443–456. doi:10.1016/j.neubiorev.2017.06.012. PMID 28666827.
- 1 2 Weng YL, An R, Cassin J, Joseph J, Mi R, Wang C, Zhong C, Jin SG, Pfeifer GP, Bellacosa A, Dong X, Hoke A, He Z, Song H, Ming GL (April 2017). "An Intrinsic Epigenetic Barrier for Functional Axon Regeneration". Neuron. 94 (2): 337–346.e6. doi:10.1016/j.neuron.2017.03.034. PMC 6007997. PMID 28426967.
- 1 2 Loh YE, Koemeter-Cox A, Finelli MJ, Shen L, Friedel RH, Zou H (February 2017). "Comprehensive mapping of 5-hydroxymethylcytosine epigenetic dynamics in axon regeneration". Epigenetics. 12 (2): 77–92. doi:10.1080/15592294.2016.1264560. PMC 5330438. PMID 27918235.