Cytochrome
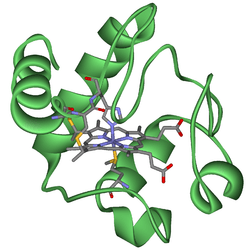
Cytochromes are heme-containing proteins. Three varieties are recognized, cytochromes a, cytochromes b, and cytochromes c. They differ with respect to the substituents on the periphery of the heme. Cytochromes function as electron relays, exploiting the switching of the iron centers from ferrous (Fe(II)) to ferric (Fe(III)).[1] In addition to cytochromes a, b, and c, many other cytochromes exist. These other cytochromes include cytochrome oxidase and cytochrome P450.
History
Cytochromes were initially described in 1884 by MacMunn as respiratory pigments (myohematin or histohematin).[2] In the 1920s, Keilin rediscovered these respiratory pigments and named them the cytochromes, or “cellular pigments”, and classified these heme proteins, on the basis of the position of their lowest energy absorption band in the reduced state, as cytochromes a (605 nm), b (~565 nm), and c (550 nm). The ultra-violet (UV) to visible spectroscopic signatures of hemes are still used to identify heme type from the reduced bis-pyridine-ligated state, i.e., the pyridine hemochrome method. Within each class, cytochrome a, b, or c, early cytochromes are numbered consecutively, e.g. cyt c, cyt c1, and cyt c2, with more recent examples designated by their reduced state R-band maximum, e.g. cyt c559.[3]
Structure and function
The heme group is a highly conjugated ring system (which allows its electrons to be very mobile) surrounding a metal ion, which readily interconverts between the oxidation states. For many cytochromes, the metal ion present is that of iron, which interconverts between Fe2+ (reduced) and Fe3+ (oxidized) states (electron-transfer processes. Cytochromes are, thus, capable of performing oxidation and reduction. Because the cytochromes (as well as other complexes) are held within membranes in an organized way, the redox reactions are carried out in the proper sequence for maximum efficiency.
In the process of oxidative phosphorylation, which is the principal energy-generating process undertaken by organisms, other membrane-bound and -soluble complexes and cofactors are involved in the chain of redox reactions, with the additional net effect that protons (H+) are transported across the mitochondrial inner membrane. The resulting transmembrane proton gradient (protonmotive force) is used to generate ATP, which is the universal chemical energy currency of life. ATP is consumed to drive cellular processes that require energy (such as synthesis of macromolecules, active transport of molecules across the membrane, and assembly of flagella).
Many scientists believe that cytochrome c has evolved at a fairly constant rate. The constant rate of change in cytochrome c may be the basis for a molecular "clock". The molecular clock can be a helpful tool in trying to determine when various organisms may have diverged from a common ancestor.[4]
Types
Several kinds of cytochrome exist and can be distinguished by spectroscopy, exact structure of the heme group, inhibitor sensitivity, and reduction potential.
Three types of cytochrome are distinguished by their prosthetic groups:
| Type | Prosthetic group |
|---|---|
| Cytochrome a | heme a |
| Cytochrome b | heme b |
| Cytochrome d | tetrapyrrolic chelate of iron[5] |
The definition of cytochrome c is not defined in terms of the heme group.[6] There is no "cytochrome e," but there is a cytochrome f, which is often considered a type of cytochrome c.[7]
In mitochondria and chloroplasts, these cytochromes are often combined in electron transport and related metabolic pathways:[8]
| Cytochromes | Combination |
|---|---|
| a and a3 | Cytochrome c oxidase ("Complex IV") with electrons delivered to complex by soluble cytochrome c (hence the name) |
| b and c1 | Coenzyme Q - cytochrome c reductase ("Complex III") |
| b6 and f | Plastoquinol—plastocyanin reductase |
A distinct family of cytochromes is the cytochrome P450 family, so named for the characteristic Soret peak formed by absorbance of light at wavelengths near 450 nm when the heme iron is reduced (with sodium dithionite) and complexed to carbon monoxide. These enzymes are primarily involved in steroidogenesis and detoxification.
References
- ↑ L., Lehninger, Albert (2000). Lehninger Principles of Biochemistry (3rd ed.). New York: Worth Publishers. ISBN 978-1572591530. OCLC 42619569.
- ↑ Mac Munn, C. A (1886). "Researches on Myohaematin and the Histohaematins". Philosophical Transactions of the Royal Society of London. 177: 267–298. doi:10.1098/rstl.1886.0007. JSTOR 109482.
- ↑ Reedy, C. J.; Gibney, B. R. (February 2004). "Heme protein assemblies". Chem Rev. 104 (2): 617–49. doi:10.1021/cr0206115. PMID 14871137.
- ↑ http://pdecandia.com/Honors%20Bio%20PDF/handout%20and%20wksts/evo%20making%20a%20molecular%20clock%20wkst.pdf
- ↑ Cytochrome+d at the US National Library of Medicine Medical Subject Headings (MeSH)
- ↑ Cytochrome+c+Group at the US National Library of Medicine Medical Subject Headings (MeSH).
- ↑ Bendall, Derek S. (2004). "The Unfinished Story of Cytochrome f". Photosynthesis Research. 80 (1–3): 265–276. doi:10.1023/b:pres.0000030454.23940.f9. ISSN 0166-8595. PMID 16328825.
- ↑ Doidge, Norman (2015). The brain’s way of healing : remarkable discoveries and recoveries from the frontiers of neuroplasticity. Penguin Group. p. 173. ISBN 978-0-698-19143-3.
External links
- Scripps Database of Metalloproteins
- Cytochromes at the US National Library of Medicine Medical Subject Headings (MeSH)