Aerosaurus
| Aerosaurus | |
|---|---|
 | |
| Life restoration of Aerosaurus wellesi | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Family: | †Varanopidae |
| Subfamily: | †Varanopinae |
| Genus: | †Aerosaurus Romer, 1937 |
| Species | |
| |
Aerosaurus (meaning "copper lizard") is an extinct genus within Varanopidae, a family of non-mammalian synapsids. It lived between 252-299 million years ago during the Early Permian in North America. The name comes from Latin aes (aeris) (combining stem: aer-) “copper” and Greek sauros “lizard,” for El Cobre Canyon (from Spanish cobre “copper”) in northern New Mexico, where the type fossil was found and the site of former copper mines. Aerosaurus was a small to medium-bodied carnivorous synapsid characterized by its recurved teeth, triangular lateral temporal fenestra, and extended teeth row. Two species are recognized: A. greenleeorum (1937) and A. wellesi (1981).[1]
History and Discovery
Aerosaurus was a taxon named by American paleontologist and biologist Alfred Romer in 1937. There has only been two specimen discoveries A. greenleeorum (1937) and A. wellesi (1981) from New Mexico. The discovery of A. wellesi came from a team led by Dr. Samuel P. Welles in 1935 from blocks of red siltstone measuring thirty square feet in the Lower Permian Abo/Cutler Formation in near the Arroyo del Aqua settlement. The blocks of siltstone would have contained two incomplete skulls of Oedaleops campi Langston, 1965, and the two nearly complete articulated skeletons of a varanopseid pelycosaur. A. greenleeorum Romer, 1937, is based on bone fragments assumed from a single individual collected by Romer in El Cobre Canyon near the Abo/Cutler Formation of the Arroyo del Agua. Cross examination of fragment holotype of A. greenleeorum Romer with the Camp Quarry skeleton with its expanded tooth-bearing parasphenoidal plate matches it to Aerosaurus taxa.[2]
Description
Aerosaurus has been recognized as a basal member of the Synapsida, a clade that ultimately produced mammals. Relative to the skull, the body was much larger and the ribs were deeper. The tail was unusually long for a synapsid. Overall Varanopseid pelycosaurs measure about one meter long with fewer maxillary and dentary teeth compared to other Varanops. In Aerosaurus, the teeth were more laterally compressed creating a strong recurved shape. Aerosaurus cranial morphology is distinguishable with its extremely long tooth row, recurved tooth, triangular lateral temporal fenestra. The teeth were so highly curved and compressed that they seem unable to penetrate flesh and the tooth row extended far behind the orbit. The lower teeth were also relatively tiny and such an arrangement suggests Aerosaurus was a carnivore. Their limbs were long, and skeleton built lightly suggests they were active and agile. A long low skull and dentition extended far back to lie below the temporal fenestra. Aerosaurus had other general features found in ‘Pelycosaurs’ like a slanted Occiput, a lateral temporal fenestra, a septomaxilla bone that contacting the nasal and a maxilla bone contacting the quadrojugal.[2]
Skull
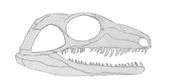
The skull is 1/4 of the precaudal length of the skeleton with a jaw articulation far back seen in Varanops. The snout is long and low with an lessen inclined occiput. The dermal bones are shallow, irregular with pits and grooves and below the orbit we find a saw-like serration that hasn't been seen in other 'pelycosaurs'. The skull roof has large round orbits and an undetermined laterally expanded prefrontal process. The postorbital is incomplete, but the postorbital and suborbital bars are thinly shaped. The pineal foramen is larger compared to non-varanopseid sphenacodonts and is positioned in the back of the skull. The lateral temporal fenestra of Aerosaurus is larger than other pelycosaurs with its trademark triangular shape. No septomaxilla is evident, but the premaxilla is fragile in comparison to the large naris. The maxilla contains specimens largest teeth and is extended posterior to the center of the triangular lateral temporal fenestra. The maxilla, which is longer compared to other 'pelycosaurs' contacts the quadratojugal in this specimen. The lacrimal is a sheet of bone that covers most of the anterior orbital margin and constricts the front of the orbit. The jugal has a unique lateral serration and narrow postorbital ramus similar to Varanops. The quadtrojugal has a slender ramus that meets the jugal and the maxilla. The nasal is also long and slender, but has extensive fragmentation, so an exact length is undetermined. The frontal is long and slender with the posterior end not extending into the margins of the orbit. The Postfrontal is small due to the size of the frontal. The squamosal is long and narrow and even more so compared to Varanops. The parietal is large with shallow grooves in the supratemporal element. The braincase is well preserved with only the cultriform process missing in all of its external surfaces. The parasphenoid is the largest part of the braincase with some overlap into the basiooccipital to form the "fenestra ovalis". The presence of the many strong, pointed, and recurved teeth is a distinguishing feature of the bottom side of the braincase. The teeth are arranged in a pattern of three groups to form a U-shaped band. There is trouble distinguishing between the basisphenoid and the parasphenoid due to the co-ossification creating a vague suture line. The basisphenoid has a large basipterygoid process that extend from the base of the cultriform process.[2]
Mandible
The mandible has a long and slender shape with grooves and pits on the skull roof. The dentary is long with a posterodorsal process that extends into the lower jaw and overlapping the surangular. The angular and prearticular bones are long with the splenial almost as long as the dentary. The coronoid was unidentifiable in this specimen. A foramen intermandibularis caudalis is on the medial surface of the mandible similar to the related Varannops and Ophiacodon species.[2]
Dentition
The skull of Aerosaurus had 16 marginal teeth, less than in Varanops (44) and the upper teeth were longer. Most of the teeth were recurved more than other 'pelycosaurs' with a triangular depression near the base. The size of each teeth varied with some teeth sized gap spaces. There largest teeth (7) were on the maxilla and decreased in length posterior to the orbit. There is additional six long recurved teeth located ventral to the postorbital bar, but the function of them are a mystery. The mandibular teeth are smaller, closely spaced, uniform in its recurved shape compared to the maxilla.[2]
Vertebrae
There are twenty-six presacral vertebrae with some missing due to erosion. Around five of the anterior vertebrae could possibly be cervical and three sacral vertebrae are present in this specimen. Overall there are at least 70 caudal vertebrae making up its long slender tail, which is greater than any other 'pelycosaur'. The neural spines are shorter with a typical atlas bearing a posterior spine to pass to the zygapophyses. The axix is about the length of the other cervical vertebrae. Each vertebrae is ossified with no visible suture between the centrum and neural arch. The distal caudal vertebrae is slender and cylinder shaped and is poorly preserved compared to the other portion of vertebrae.[2]
Ribs
Presacral vertebrae have a slender double-headed ribs, but which rib is the longest is undetermined because of the articulation of the fossil. Some dorsal ribs are distally expanded. The Gastralia is seen in a cluster of tiny rods in some articulated ribs. Gastralia are narrow plates in other 'pelycosaurs' compared to Aerosaurus.[2]
Appendicular Skeleton
The pectoral girdle is preserved in a medial surface with the left scapula partially exposed. The clavicle is also preserved, but no cleithrum is identifiable in this specimen. The shoulder girdle similar to Varanops with a curved scapulocoracoid. The clavicle is expanded anteroposteriorly. The T-shaped interclavicle is similar to Ophiacodon and is well preserved. The pelvic girdle is not entirely exposed in this specimen. The neck of the iliac blade is narrower compared to A. greenleeorum, but the ischium is longer and ossified. The pubis is not exposed well enough to be described. The left side of the limbs are well preserved with the humerus and femur in similar length to the lower legs. Allowing for a high posture and long strides for a predaceous mode of life. The ulna is straight, thick with a well developed olecranon. The left manus is assumed to be complete with what is visible in the fossil. Terminal phalanges have short recurved claws with a phalangeal formula of 2:3:4:5:3. The femoras are straight, thick with ends that aren't enlarged. The tibia is big and bowed shaped. The astragulus is short and square-cut without a long proximal neck.[2]
Paleobiology
A. wellesi had a humerus and femur in similar length to the lower legs, which allowed it to have a high posture and long strides for a predaceous or carnivorous type of life. The ossified feet and exceedingly long tail of Aerosaurus suggests an agile terrestrial carnivore[2] ‘Pelycosaurs’ represent first group to dominate the terrestrial fauna and achieve large body size at a time where the supercontinent Pangea existed. The discovery of Pyozia mesenensis from the Middle Permian suggests there was an adaptation to ecological and climate changes that occurs into the late Paleozoic. Varanopid tended to be small to medium-sized basal 'pelycosaurs' that were carnivores and coexisted with herbivory animals in this landscape. The P. mesenensis discovery is just an increasing trend of a “mammalness” trait during the introduction of herbivory prey that appears during the Permian.[3]
Thermoregulation
Members of 'pelycosaurs' were most likely ectothermic with some theories with Edaphosauridae and Dimetrodon using their sail structure to preserve heat. Ectothermic animals like reptiles and 'pelycosaurs' needed to maintain a level of body temperature versus the external environment in order to survive. It is estimated that the 'pelycosaur' would need to stay within the range of 24 to 35 degrees Celsius similar to modern reptiles. This is backed up by the location of 'pelycosaur discoveries in the regions of warmer temperatures in the supercontinent of Pangea in now known localities in New Mexico, Texas and Oklahoma to name a few. A change in color, respiration, blood flow and metabolism are some of the ways reptiles today regulate their body temperature and could be some of the ways 'pelycosaur' could evolve to expand their niche.[4]
Paleoenvironment
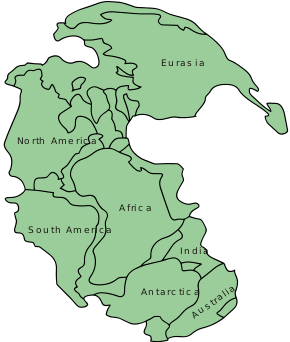
The 'pelycosaurs' and Aerosaurus would have lived in the super continent Pangaea. The northern and southern hemisphere during the Early Permian was hot and arid with evaporite deposits in North America and Europe. The southernmost and Northernmost continents of Pangaea had a cold and temperate climate. The equator of Pangaea would have a tropical and humid climate. The flora would change from the tropical forests of the Carboniferous to be replaced by seed-bearing plants like conifers during the Early Permian. The most discoveries from the Late Carboniferous and Early Permian tetrapods were in the Northern Hemisphere. Discoveries of fossil tetrapods from the Middle and Late Permian were from South Africa and Russia creating a temporal gap. A temporal gap between the fossil records of the Early and Middle Permian or the Olson's Gap, after Everett C. Olson, a well known vertebrate paleontologist was closed with redating Permian fossil beds and newer fossil discoveries. Aerosaurus would be found in red siltstone in the Abo/Cutler Formation in New Mexico, so it existed within an open and arid environment.[5]
Classification
Aerosaurus belongs in the extinct family Varanopidae also called a "pelycosaurs" or "mammal-like" reptile, they became extincted around the end of the Permian. Within the Varanopidae family there are the subfamilies Mycterosaurinae and the Varanopinae, the Varanopinae is where Aerosaurus belongs in this phylogeny. Elliotsmithia longiceps is a closely related subfamily to Aerosaurus and is grouped in Mycterosaurinae. The sister taxa of Varanopidae is the Ophiacontidae family.[6]
|
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Pelycosaurs
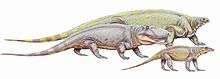
The 'pelycosaurs' existed during the Early Permian, Middle Permian, Late Permian, and to the Early Triassic. 'Mammal-like Reptiles' or pelycosaur is based on the morphology of their dentary and marginal teeth.[7] However, there is a significant negative correlation between skeletal completeness and the diversity of 'pelycosaurs' suggests many species were named using poor specimens. The majority of pelycosaur species named during the 1930s to the 1960 were based on size and location rather than character based morphology to suggest that there is a great deal of missing fossil record.[8] Basal synapsids or ‘pelycosaurs’ form the beginning portion of the mammalian stem lineage from ‘reptile-like’ to the more mammal derived traits we find in therapsids. ‘Pelycosaurs dominated the terrestrial ecosystem from the Carboniferous to the Early Permian creating animals 3-6.5 meters long, herbivores, macropredators and a diversity of smaller bodied vertebrate. Postcranial data and a full dataset suggests that the clade Ophiacodontidae and Varanopidae is the most basal clade.[9]
Varanopidae

The genus Varanopidae has eight genres and has one of the longest fossil record starting from the Carboniferous to the Middle Permian. Varanopids are small to medium-sized synapsids that has been discovered throughout the super continent Pangea. Varanopids are found in formerly areas of North America, Russia, Europe, and South Africa.[6] The authors Romer and Price (1940) discussed the original positioning of Varanopidae within Synapsida and considered them as the suborder Sphenacodontia. Recent phylogenetic analyses has place Varanopidae as a basal member of Synapsida and due to their positioning, a better understanding of the morphology and phylogeny of varanopids is needed for synapsid evolution. The phylogeny of varanopids should be based mostly on cranial morphology.[10][11] The atlas−axis complex can be described with little effort with variation of this structure within a small clade. Varanopids, members of synapsid predators have well preserved atlas−axes permitting a descriptions and examination of morphological variation between taxon. The size of the transverse processes on the axis and the shape of the axial neural spine can be variable. For the small mycterosaurine varanopids, they have a small transverse processes that point posteroventrally, and the axial spine is dorsoventrally short, with a flattened dorsal margin in lateral view. The larger varanodontine varanopids have large transverse processes with a broad base, and a much taller axial spine with a rounded dorsal margin in lateral view. Using outgroup comparisons, the morphology of the transverse processes is considered a derived trait in varanodontines, while in mycterosaurines the morphology of the axial spine is the derived trait.[12]
Herbivory
An extinction event would influence the evolution of the pelcosaurian synapsids during the Kasimovian. For Varanopidae, it would cause a transition to a more open and arid habitat decreasing the diversification of synapsids. Synapsid herbivore evolution is suggested to occur during the late Carboniferous and Permian. Edaphosauridae and Caseidae, two pelycosaurian synapsids are suggested to be a herbivorous species. Edaphosauridae would have a large tooth plate and lingual surface of the lower jaw to crush vegetation (Modesto, 1995), Caseidae would use a spatulate teeth for shredding of vegetation.[13][14]
See also
References
- ↑ Romer, A. S. (1937). "New Genera and Species of Pelycosaurian Reptiles" (PDF). Proceedings of the New England Zoölogical Club. 16: 90–96.
- 1 2 3 4 5 6 7 8 9 Langston, W.; Reisz, R. (1981). "Aerosaurus wellesi, new species, a varanopseid mammal-like reptile (Synapsida : Pelycosauria) from the Lower Permian of New Mexico". Journal of Vertebrate Paleontology. 1 (1): 73–96. JSTOR 4522838.
- ↑ Anderson, J.; Reisz, R. (2004). "Pyozia mesenensis, a new, small varanopid (Synapsida, Eupelycosauria) from Russia: "pelycosaur" diversity in the Middle Permian". Journal of Vertebrate Paleontology. 24 (1): 173–179. doi:10.1671/1940-13. JSTOR 4524702.
- ↑ Florides, G. A.; Wrobel, L. C.; Kalogirou, S. A.; Tassou, S. (1999). "A thermal model for reptiles and pelycosaurs". Journal of Thermal Biology. 24 (1): 1–13. doi:10.1016/S0306-4565(98)00032-1.
- ↑ Benton, M.; Sibbick, J. (2015). Vertebrate palaeontology (4 ed.). West Sussex, England: John Wiley & Sons. ISBN 978-1-118-40755-4.
- 1 2 Modesto, S.; Sidor, C.; Rubidge, B.; Welman, J. (2001). "A Second Varanopseid Skull from the Upper Permian of South Africa: Implications for Late Permian 'Pelycosaur' Evolution". Lethaia. 34 (4): 249–259. doi:10.1111/j.1502-3931.2001.tb00053.x.
- ↑ Modesto, S.; Smith, R.; Campione, N.; Reisz, R. (2011). "The Last "Pelycosaur": A Varanopid Synapsid from the Pristerognathus Assemblage Zone, Middle Permian of South Africa". Naturwissenschaften. 98 (12): 1027–1034. doi:10.1007/s00114-011-0856-2. PMID 22009069.
- ↑ Brocklehurst, N.; Frobisch, J.; Froebisch, N. (2014). "Current and Historical Perspectives on the Completeness of the Fossil Record of Pelycosaurian-grade Synapsids". Palaeogeography, Palaeoclimatology, Palaeoecology. 399: 114–126. doi:10.1016/j.palaeo.2014.02.006.
- ↑ Benson, R. (2011). "Interrelationships of Basal Synapsids: Cranial and Postcranial Morphological Partitions Suggest Different Topologies". Journal of Systematic Palaeontology. 10 (4): 601–624. doi:10.1080/14772019.2011.631042.
- ↑ Maddin, H. C.; Evans, D. C.; Reisz, R. R. (2006). "An Early Permian Varanodontine Varanopid (Synapsida: Eupelycosauria) from the Richards Spurs Locality, Oklahoma". Journal of Vertebrate Paleontology. 26 (4): 957–966. doi:10.1671/0272-4634(2006)26[957:AEPVVS]2.0.CO;2. JSTOR 4524646.
- ↑ Campione, N.; Reisz, R. (2010). "Varanops brevirostris (Eupelycosauria: Varanopidae) from the Lower Permian of Texas, with Discussion of Varanopid Morphology and Interrelationships". Journal of Vertebrate Paleontology. 30 (3): 724–746. doi:10.1080/02724631003762914.
- ↑ Campione, N. E.; Reisz, R. R. (2011). "Morphology and Evolutionary Significance of the Atlas-axis Complex in Varanopid Synapsids" (PDF). Acta Palaeontologica Polonica. 56 (4): 739–748. doi:10.4202/app.2010.0071.
- ↑ Sues, H. D.; Reisz, R. R. (1998). "Origins and Early Evolution of Herbivory in Tetrapods". Trends in Ecology and Evolution. 13 (4): 141–145. doi:10.1016/S0169-5347(97)01257-3. PMID 21238234.
- ↑ Brocklehurst, N.; Kammerer, C.; Fröbisch, J. (2013). "The Early Evolution of Synapsids, and the Influence of Sampling on their Fossil Record". Paleobiology (39): 470–490. doi:10.1666/12049. JSTOR 42004210.
External links
![]()
| Ophiacodontidae |  | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Varanopidae |
| ||||||||||||
| Caseasauria |
| ||||||||||||
| Edaphosauridae | |||||||||||||
| Sphenacodontia |
| ||||||||||||
| Others | |||||||||||||
| Related categories |
| ||||||||||||