Membrane protein
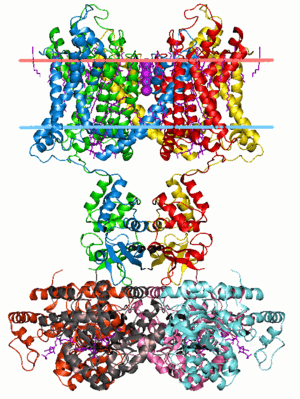
Membrane proteins are proteins that interact with, or are part of, biological membranes. They include integral membrane proteins that are permanently anchored or part of the membrane and peripheral membrane proteins that are only temporarily attached to the lipid bilayer or to other integral proteins.[1][2] The integral membrane proteins are classified as transmembrane proteins that span across the membrane and integral monotopic proteins that are attached to only one side of the membrane. Membrane proteins are a common type of proteins along with soluble globular proteins, fibrous proteins, and disordered proteins.[3] They are targets of over 50% of all modern medicinal drugs.[4] It is estimated that 20–30% of all genes in most genomes encode membrane proteins.[5][6]
Compared to other classes of proteins, the determination of membrane protein structures has remained a challenge in large part due to the difficulty in establishing experimental conditions where the correct conformation of the protein in isolation from its native environment is preserved.[6]
Function
Membrane proteins perform a variety of functions vital to the survival of organisms:[7]
- Membrane receptor proteins relay signals between the cell's internal and external environments.
- Transport proteins move molecules and ions across the membrane. They can be categorized according to the Transporter Classification database.
- Membrane enzymes may have many activities, such as oxidoreductase, transferase or hydrolase.[8]
- Cell adhesion molecules allow cells to identify each other and interact. For example, proteins involved in immune response.
Integral membrane proteins
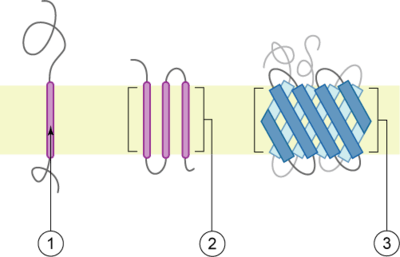
The membrane is represented in light-brown.
Integral membrane proteins are permanently attached to the membrane. Such proteins can be separated from the biological membranes only using detergents, nonpolar solvents, or sometimes denaturing agents. One such example of this type of protein which has not been functionally characterized yet is SMIM23. They can be classified according to their relationship with the bilayer:
- Integral polytopic proteins are transmembrane proteins that span across the membrane more than once. These proteins may have different transmembrane topology.[9][10] These proteins have one of two structural architectures:
- helix bundle proteins, which are present in all types of biological membranes;
- beta barrel proteins, which are found only in outer membranes of Gram-negative bacteria, and outer membranes of mitochondria and chloroplasts.[11]
- Bitopic proteins are transmembrane proteins that span across the membrane only once. Transmembrane helices from these proteins have significantly different amino acid distributions to transmembrane helices from polytopic proteins.[12]
- Integral monotopic proteins are integral membrane proteins that are attached to only one side of the membrane and do not span the whole way across.
Peripheral membrane proteins
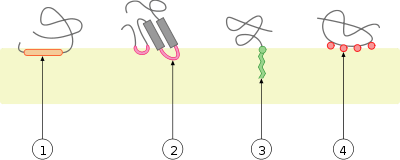
Peripheral membrane proteins are temporarily attached either to the lipid bilayer or to integral proteins by a combination of hydrophobic, electrostatic, and other non-covalent interactions. Peripheral proteins dissociate following treatment with a polar reagent, such as a solution with an elevated pH or high salt concentrations.
Integral and peripheral proteins may be post-translationally modified, with added fatty acid or prenyl chains, or GPI (glycosylphosphatidylinositol), which may be anchored in the lipid bilayer.
Polypeptide toxins
Polypeptide toxins and many antibacterial peptides, such as colicins or hemolysins, and certain proteins involved in apoptosis, are sometimes considered a separate category. These proteins are water-soluble but can aggregate and associate irreversibly with the lipid bilayer and become reversibly or irreversibly membrane-associated.
In genomes
A large fraction of all proteins are thought to be membrane proteins. For instance, about 1000 of the ~4200 proteins of E. coli are thought to be membrane proteins.[13] The membrane localization has been confirmed for more than 600 of them experimentally.[13] The localization of proteins in membranes can be predicted very reliably using hydrophobicity analyses of protein sequences, i.e. the localization of hydrophobic amino acid sequences.
See also
References
- ↑ Johnson JE, Cornell RB (1999). "Amphitropic proteins: regulation by reversible membrane interactions (review)". Mol. Membr. Biol. 16 (3): 217–235. doi:10.1080/096876899294544. PMID 10503244.
- ↑ Alenghat, Francis J.; Golan, David E. (2013). "Membrane Protein Dynamics and Functional Implications in Mammalian Cells". Current Topics in Membranes. Current Topics in Membranes. 72: 89–120. doi:10.1016/b978-0-12-417027-8.00003-9. ISBN 9780124170278. PMC 4193470. PMID 24210428.
- ↑ Andreeva, A (2014). "SCOP2 prototype: a new approach to protein structure mining". Nucleic Acids Res. 42 (Database issue): D310–4. doi:10.1093/nar/gkt1242. PMC 3964979. PMID 24293656.
- ↑ Overington JP, Al-Lazikani B, Hopkins AL (December 2006). "How many drug targets are there?". Nat Rev Drug Discov. 5 (12): 993–6. doi:10.1038/nrd2199. PMID 17139284.
- ↑ Krogh, A.; Larsson, B. R.; Von Heijne, G.; Sonnhammer, E. L. L. (2001). "Predicting transmembrane protein topology with a hidden markov model: Application to complete genomes". Journal of Molecular Biology. 305 (3): 567–580. doi:10.1006/jmbi.2000.4315. PMID 11152613.
- 1 2 Liszewski, Kathy (1 October 2015). "Dissecting the Structure of Membrane Proteins". Genetic Engineering & Biotechnology News (paper). 35 (17): 1, 14, 16&ndash, 17. doi:10.1089/gen.35.17.02. (subscription required)
- ↑ Almén, M.; Nordström, K. J.; Fredriksson, R.; Schiöth, H. B. (2009). "Mapping the human membrane proteome: A majority of the human membrane proteins can be classified according to function and evolutionary origin". BMC Biology. 7: 50. doi:10.1186/1741-7007-7-50. PMC 2739160. PMID 19678920.
- ↑ Yibin, Lin (2013). "The substitution of Arg149 with Cys fixes the melibiose transporter in an inward-open conformation". Biochimica et Biophysica Acta. 1828 (8): 1690. doi:10.1016/j.bbamem.2013.03.003.
- ↑ Von Heijne, G. (2006). "Membrane-protein topology". Nature Reviews Molecular Cell Biology. 7 (12): 909–918. doi:10.1038/nrm2063. PMID 17139331.
- ↑ Gerald Karp (2009). Cell and Molecular Biology: Concepts and Experiments. John Wiley and Sons. pp. 128–. ISBN 978-0-470-48337-4. Retrieved 13 November 2010.
- ↑ Selkrig J, Leyton DL, Webb CT, Lithgow T (2014). "Assembly of β-barrel proteins into bacterial outer membranes". Biochimica et Biophysica Acta. 1843 (8): 1542–50. doi:10.1016/j.bbamcr.2013.10.009. PMID 24135059.
- ↑ Baker, James Alexander; Wong, Wing-Cheong; Eisenhaber, Birgit; Warwicker, Jim; Eisenhaber, Frank (2017). "Charged residues next to transmembrane regions revisited: "Positive-inside rule" is complemented by the "negative inside depletion/outside enrichment rule"". BMC Biology. 15 (1): 66. doi:10.1186/s12915-017-0404-4.
- 1 2 Daley, D. O.; Rapp, M; Granseth, E; Melén, K; Drew, D; von Heijne, G (2005). "Global topology analysis of the Escherichia coli inner membrane proteome". Science. 308 (5726): 1321–3. doi:10.1126/science.1109730. PMID 15919996.
External links
| Look up membrane protein in Wiktionary, the free dictionary. |
| Wikimedia Commons has media related to Membrane proteins. |
Organizations
Membrane protein databases
- TCDB - Transporter Classification database, a comprehensive classification of transmembrane transporter proteins
- Orientations of Proteins in Membranes (OPM) database 3D structures of integral and peripheral membrane proteins arranged in the lipid bilayer
- Protein Data Bank of Transmembrane Proteins 3D models of all transmembrane proteins currently in PDB. Approximate positions of membrane boundary planes were calculated for each PDB entry.
- TransportDB Genomics-oriented database of transporters from TIGR
- Membrane PDB Database of 3D structures of integral membrane proteins and hydrophobic peptides with an emphasis on crystallization conditions
- Mpstruc database List of transmembrane proteins of known 3D structure. A curated list of transmembrane proteins from the Protein Data Bank
- MemProtMD is a database of around 3000 intrinsic membrane protein structures
- Membranome database provides information about bitopic proteins from several model organisms