Lactase persistence
Lactase persistence is the continued activity of the lactase enzyme in adulthood. Since lactase's only function is the digestion of lactose in milk, in most mammal species, the activity of the enzyme is dramatically reduced after weaning.[1] In some human populations, though, lactase persistence has recently evolved[2] as an adaptation to the consumption of nonhuman milk and dairy products beyond infancy. The majority of people around the world remain lactase nonpersistent,[1] and consequently are affected by varying degrees of lactose intolerance as adults. However, not all genetically lactase nonpersistent individuals are noticeably lactose intolerant, and not all lactose intolerant individuals have the lactase nonpersistence genotype.
Global spread of the lactase persistence phenotype
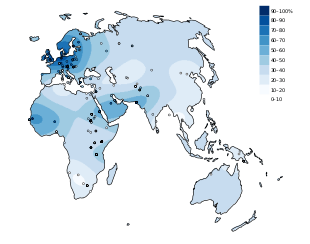
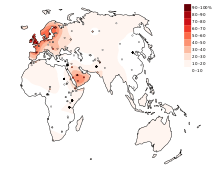
The distribution of the lactase persistence phenotype, or the ability to digest lactose into adulthood, is not homogeneous in the world. Lactase persistence-frequencies are highly variable. In Europe, the distribution of the lactase persistence phenotype is clinal, with frequencies ranging from 15–54% in the south-east to 89–96% in the north-west.[3] For example, only 17% of Greeks and 14% of Sardinians are predicted to possess this phenotype, while around 80% of Finns and Hungarians and 100% of Irish people are predicted to be lactase persistent.[4]
High frequencies of lactase persistence are also found in some places in Sub-Saharan Africa and in the Middle East. But the most common situation is intermediate to low lactase persistence: intermediate (11 to 32%) in Central Asia,[5] low (<=5%) in Native Americans, East Asians, most Chinese populations[2] and some African populations.[3][6][5]
In Africa, the distribution of lactase persistence is "patchy":[7][8][3] high variations of frequency are observed in neighbouring populations, for example between Beja and Nilotes from Sudan.[9] This makes the study of lactase persistence distribution more difficult.[6] High percentages of lactase persistence phenotype are found in traditionally pastoralist populations like Fulani and Bedouins.[3][10] It is absent in the Bantu of South Africa.[2]
Genetics
Multiple studies indicate that the presence of the two phenotypes "lactase persistent" (derived phenotype) and "lactase nonpersistent (hypolactasia)" is genetically programmed, and that lactase persistence is not necessarily conditioned by the consumption of lactose after the suckling period.[11][12]
The lactase persistent phenotype involves high mRNA expression, high lactase activity, and thus the ability to digest lactose, while the lactase nonpersistent phenotype involves low mRNA expression and low lactase activity.[13] The enzyme lactase is encoded by the gene LCT.[11]
Hypolactasia is known to be recessively and autosomally inherited, which means that individuals with the nonpersistent phenotype are homozygous and received the two copies of a low lactase-activity allele (the ancestral allele) from their parents, who may be homozygous or at least heterozygous for the allele.[11] Only one high-activity allele is required to be lactase persistent.[11][12] Lactase persistence behaves as a dominant trait because half levels of lactase activity are sufficient to show significant digestion of lactose.[1] Cis-acting transcriptional silence of the lactase gene is responsible for the hypolactasia phenotype.[11][12] Furthermore, studies show that only eight cases were found where the parents of a child with lactase persistence were both hypolactasic.[1] While a variety of genetic, as well as nutritional, factors determine lactase expression, no evidence has been found for adaptive alteration of lactase expression within an individual in response to changes in lactose consumption levels.[1] The two distinct phenotypes of hypolactasia are: Phenotype I, characterized by reduced synthesis of precursor LPH, and phenotype II, associated with ample precursor synthesis, but reduced conversion of the protein to its mature molecular form.[14] The lactase enzyme has two active sites which break down lactose. The first is at Glu1273 and the second is at Glu1749, which separately break down lactose into two separate kinds of molecules.[1]
At least six mutations (single-nucleotide polymorphisms – SNPs) have been associated with lactase expression.[15] They are all located in a region of the gene MCM6 upstream of LCT. This region is considered as an enhancer region for the transcription of LCT.[16][17][18] The first identified genetic variant associated with lactase persistence is C/T*−13910.[19] The ancestral allele is C and the derived allele – associated with lactase persistence – is T. In the same study, another variant was found to also correlate with the phenotype in most of the cases: G*/A-22018.
Other alleles associated with lactase persistence have been identified: G/C*-14010,[10] C/G*-13907,[10][8][20] and T/G*-13915.[21] This variant is described as part of a compound allele with T/C*3712 in.[18] These three variants are widespread in some populations. Rare variants were reported in a few studies, like the G/A*14107 in the Xhosa[22] and the Fulani (from Mali);[15] the C/T*13906 in the Fulani (from Mali).[15]
Lactase-persistence alleles vary in their geographic distributions. Within European and populations of European ancestry, they are almost entirely correlated with the presence of the −13,910 C/T mutation in the enhancer region of the lactase gene (LCT).
This differs from lactase persistence allelic distributions in the rest of the world, particularly in Africa and in the Middle East, where several alleles coexist.
The T/G*-13915 allele is found mostly in populations from East and North Africa and the Middle East. The allele G/C*-14010 was identified in East Africa.[23] The C/G*13907 allele was described in Sudan and Ethiopia.[10][20][24] The "European" allele T*13910 allele is also found in some populations from Africa, including the Fulani (from Mali,[15] Sudan,[25] and Cameroon[20]) and the Khoe from South Africa.[26][27] This allele has also been found in Central Asia.[5]
It is not known how exactly the different variants described above regulate LCT expression. None of the mutations so far identified have been shown to be exclusively causal for lactase persistence, and it is possible that there are more alleles to be discovered.[28] If we focus on the "European variant", the position −13910 has an enhancer function on the lactase promoter (the promoter facilitates the transcription of the LCT gene). T−13910 is a greater enhancer than C−13910, so this mutation is thought to be responsible for the differences in lactase expression,[29] although not enough evidence is found to prove that lactase persistence is only caused by C−13910→T−13910.[11]
In addition, it was shown in one study involving a Finnish population that the lactase gene has a higher expression when G−22018 is combined with T-13910.[11]
Evolutionary advantages
Lactase persistence is a text-book example of natural selection in humans: it has been reported to present stronger selection pressure than any other known human gene.[11] However, the specific reasons as to why lactase persistence confers a selective advantage "remain open to speculation".[30]
Several pieces of evidence for positive selection acting at the T*-13910 allele were given: it is located in a stretch of homozygosity of c. 1 Mb;[31] the strength of selection is similar to that estimated for the resistance to malaria.[2] Haplotype inferences were performed on data from Central Asia populations; selection was detected there as well – though less strong than in European populations.[5] Thus, even if T*13910 may not be causative for lactase persistence, it was selected during human evolutionary history.
The other variants were also proved to be under selection. The C*-14010 allele is located on a particularly long stretch of homozygosity (> 2 Mb).[10]
The compound allele G*-13915 & C*-3712 was proved to be located on a long stretch of homozygosity (1.1 Mb[10] to 1.3 Mb[18]).
The ability to digest lactose is not an evolutionary novelty in human populations. Nearly all mammals begin life with the ability to digest lactose. This trait is advantageous during the infant stage, because milk serves as the primary source for nutrition. As weaning occurs, and other foods enter the diet, milk is no longer consumed. As a result, the ability to digest lactose no longer provides a distinct fitness advantage.[32] This is evident in examining the mammalian lactase gene (LCT), which expression decreases after the weaning stage, resulting in a lowered production of lactase enzymes.[32] When these enzymes are produced in low quantities, lactose non-persistence (LNP) results.[25]
The ability to digest fresh milk through adulthood is genetically coded for by different variants which are located upstream of the LCT gene and which differ among populations. Those variants are found at very high frequencies in some populations and show signatures of selection. Several hypotheses have been proposed to explain why the lactase persistence phenotype has been positively selected.
Gene-culture coevolution hypothesis
The gene–culture coevolution hypothesis of the positive selection of the lactase persistence phenotype is based on the observation that pastoralist populations often present high levels of lactase persistence. According to this hypothesis, the reason of selection is the nutritional advantage of being lactase persistent.[2][10]
Individuals who expressed lactase-persistent phenotypes would have had a significant advantage in nutritional acquisition.[25] This is especially true for societies in which the domestication of milk-producing animals and pastoralism became a main way of life.
The combination of pastoralism and lactase persistence genes would have allowed individuals the advantage of niche construction, meaning they would have had less competition for resources by deriving a secondary food source, milk.[33] Milk as a nutrition source may have been more advantageous than meat, as its rate of renewal is significantly faster. Rather than having to raise and slaughter animals, one cow or goat could repeatedly serve as a resource with fewer time and energy constraints. The competitive advantage conferred on lactose-tolerant individuals would have given rise to strong selective pressures for this genotype, especially in times of starvation and famine, which in turn gave rise to higher frequencies in lactase persistence within the populations.
By contrast, for societies which did not engage in pastoral behaviors, no selective advantage exists for lactase persistence. Mutations which may have developed allelic variations which code for lactase production into adulthood are simply neutral mutations. They seemingly confer no fitness benefit to individuals. As a result, no selection has perpetuated the spread of these allelic variants, and the lactase persistence genotype and phenotype remains rare.[1] For example, in East Asia, historical sources also attest that the Chinese did not consume milk, whereas the nomads who lived on the borders did. This reflects modern distributions of intolerance. China is particularly notable as a place of poor tolerance, whereas in Mongolia and the Asian steppes, milk and dairy products are a main nutrition source. The nomads also make an alcoholic beverage, called airag or kumis, from mare's milk, although the fermentation process reduces the amount of lactose present.
Two scenarios have been proposed for the gene–culture coevolution hypothesis: either lactase persistence developed and was selected after the onset of pastoralist practices (culture-historical hypothesis); or pastoralism spread only in populations where lactase persistence was already at high frequencies (reverse-cause hypothesis). There are exceptions to the hypothesis like the hunter-gatherers Hadza (Tanzania) with a prevalence of lactase persistence phenotype of 50%.[10]
Benefits of being lactase persistent in adulthood
The consumption of lactose has been shown to benefit humans with lactase persistence through adulthood. For example, the 2009 British Women's Heart and Health Study[34] investigated the effects on women's health of the alleles that coded for lactase persistence. Where the C allele indicated lactase nonpersistence and the T allele indicated lactase persistence, the study found that women who were homozygous for the C allele exhibited worse health than women with a C and a T allele and women with two T alleles. Women who were CC reported more hip and wrist fractures, more osteoporosis, and more cataracts than the other groups.[35] They also were on average 4–6 mm shorter than the other women, as well as slightly lighter in weight.[35] In addition, factors such as metabolic traits, socioeconomic status, lifestyle, and fertility were found to be unrelated to the findings, thus it can be concluded that the lactase persistence benefited the health of these women who consumed dairy products and exhibited lactase persistence.
Calcium absorption hypothesis
Another possibility is the calcium absorption hypothesis.[8][35] Lactose favors the intestinal absorption of calcium: it helps maintaining it in a soluble form. This can be advantageous in regions of low sunlight exposure where Vitamin D, necessary for the transport of calcium, is a limiting factor. Indeed, the UVB are a source of Vitamin D.
The correlation between lactase persistence frequencies and latitude in 33 populations in Europe was found to be positive and significant, while the correlation between lactase persistence and longitude was not, suggesting that high levels of lactose assimilation were indeed useful in areas of low sunlight in northern Europe.[36]
Increased calcium absorption helps to prevent rickets and osteomalacia.[1]
Arid climate hypothesis
A hypothesis specific to arid climate was proposed:[37] here, milk is not only a source of nutrients, but also a source of fluid, which could be particularly advantageous during epidemics of gastrointestinal diseases like cholera (where water is contaminated).
Lactase persistence and malaria resistance
One study suggested that lactase persistence was selected for parallel to malaria resistance in the Fulani from Mali.[15] Proposed mechanisms are: nutritional advantage of milk; low content of p-aminobenzoic acid compared to non-milk diets; intake of immunmodulators contained in milk.
Lactase non-persistence in milk reliant populations
Although the selective advantages of lactase persistence have been discussed, there have been studies of ethnic groups whose populations, despite relying heavily on milk consumption, currently have a low frequency of lactase persistence.[38] A study of 303 individuals from the Beja tribe and 282 individuals from various Nilotic tribes in Sudan discovered a sharp difference between the distribution of lactase phenotypes of the two populations. Lactase persistence was determined with hydrogen breath tests. The frequency of lactose malabsorbers was 18.4% in members of Beja tribes over the age of 30, and 73.3% in members of Nilotic tribes over the age of 30.[39]
Evolutionary history
According to the gene-culture coevolution hypothesis, the ability to digest lactose into adulthood (lactase persistence) became advantageous to humans after the invention of animal husbandry and the domestication of animal species that could provide a consistent source of milk. Hunter-gatherer populations before the Neolithic revolution were overwhelmingly lactose intolerant,[40][41] as are modern hunter-gatherers. Genetic studies suggest that the oldest mutations associated with lactase persistence only reached appreciable levels in human populations in the last 10,000 years.[42][2] This correlates with the beginning of animal domestication, which occurred during the Neolithic transition. Therefore, lactase persistence is often cited as an example of both recent human evolution[10] and, as lactase persistence is a genetic trait but animal husbandry a cultural trait, gene-culture coevolution in the mutual human-animal symbiosis initiated with the advent of agriculture.[43]
Depending on the populations, one or the other hypothesis for the selective advantage of lactase persistence is more relevant: In Northern Europe, the calcium absorption hypothesis might be one of the factors leading to the strong selection coefficients,[44] whereas in African populations, where vitamin D deficiency is not as much of an issue, the spread of the allele is most closely correlated with the added calories and nutrition from pastoralism.[2]
Several genetic markers for lactase persistence have been identified, and these show that lactase persistence has multiple origins in different parts of the world (i.e. it is an example of convergent evolution). In particular, it has been hypothesized[45] that the T*13910 variant appeared at least twice independently. Indeed, it is observed on two different haplotypes: H98, the more common (among others in the Finnish and in the Fulani); and H8 H12, related to geographically restricted populations. The common version is relatively older. The H98 variant – most common among Europeans – is estimated to have risen to significant frequencies about 7,500 years ago in the central Balkans and Central Europe, a place and time roughly corresponding to the archaeological Linear Pottery culture and Starčevo cultures.
The T*13910 variant is also found in North Africans. Thus it probably originated earlier than 7500 ya, in the Near East, but the earliest farmers did not have high levels of lactase persistence and did not consume significant amounts of unprocessed milk.[46]
Some hypotheses regarding the evolutionary history of lactase persistence in given regions of the world are described below.
Europe
Concerning Europe, the model proposed for the spread of lactase persistence combines selection and demographic processes.[34][23][3][8] Some studies used modelling approaches to investigate the role of genetic drift.[3] According to some models, the spread of lactase persistence in Europe can be attributed primarily to a form of genetic drift.[34] Evidence can also come from other fields, for example written historical records: Roman authors recorded that the people of northern Europe, particularly Britain and Germany, drank unprocessed milk. This corresponds very closely with modern European distributions of lactose intolerance, where the people of Britain, Germany, and Scandinavia have a high tolerance, and those of southern Europe, especially Italy, have a lower tolerance.
A genome-wide scan for selection using DNA gathered from 230 ancient West Eurasians who lived between 6500 and 300 BCE found that the earliest appearance of the allele responsible for lactase persistence occurred in an individual who lived in central Europe between 2450 and 2140 BCE.[47]
Central Asia
In Central Asia, the causal polymorphism for lactase persistence is the same as in Europe (T*13910, rs4988235), suggesting genetic diffusion between the two geographical regions.[5]
It is indicated that the allele responsible for lactase persistence (T*13910) may have arisen in Central Asia, based on the higher frequency of lactase persistence among Kazakhs who have the lowest proportion of "western" gene pool inferred from admixture analysis from autosomal microsatellite data.[5] This, in turn, could also be an indirect genetic proof of early domestication of horses for milk products as recently attested from archaeological remains.[5][48] In Kazakhs, traditionally herders, lactase persistence frequency is estimated to 25–32%, of which only 40.2% have symptoms and 85–92% of the individuals are carriers of the T*13910 allele.[5]
Africa
The situation is more complex in Africa, where all five main lactase persistence variants are found.[49][10][50][26]
The presence of T*13910 alleles among the Khoe pastoralists is ascribed to gene flow from Europe. However, The presence of other alleles signals gene flow from East Africa.[50]
It has been hypothesized that the G*13915 variant dispersed from the Middle East,[49] in association with the domestication of the Arabian camel.[18]
The G-14009 mutation is based in Ethiopia.[51]
The G*13907 variant is concentrated among Afroasiatic speakers in Northeast Africa.[49]
The C*14010 allele is today most common among pastoralist groups inhabiting eastern Africa, from where it is thought to have spread along with pastoralism into parts of southern Africa.[49][50][26] It is also found at high frequencies among Tuscans.[52] Ultimately, the C*14010 lactase persistence variant is believed to have arrived from the Sahara in areas that were previously inhabited by Afroasiatic-speaking populations. This was deduced from the existence of animal husbandry- and milking-related loanwords of Afroasiatic origin in various Nilo-Saharan and Niger-Congo languages, as well as from the earliest appearance of processed milk lipids on ceramics which were found at the Tadrart Acacus archaeological site in Libya (radiocarbon-dated to c. 7,500 BP, close to the estimated age of the C*14010 mutation).[53]
The evolutionary processes driving the rapid spread of lactase persistence in some populations are not known.[1] Among some populations inhabiting East Africa, lactase persistence has gone from negligible to near-ubiquitous frequencies in just 3000 years, suggesting a very strong selective pressure.[10] Some studies also proposed that selection for lactase persistence is not as strong as supposed (soft selective sweep), and that its strength varies a lot depending on particular environmental conditions.[8]
In nonhumans
Lactose malabsorption is typical for adult mammals, and lactase persistence is a phenomenon likely linked to human interactions in the form of dairying. Most mammals lose the ability to digest lactose once they are old enough to find their own source of nourishment away from their mothers.[54] After weaning, or the transition from being milk-fed to consuming other types of food, their ability to produce lactase naturally diminishes as it is no longer needed. For example, in the time a piglet in one study aged from five to 18 days, it lost 67% of its lactose absorption ability.[55] While nearly all humans can normally digest lactose for the first 5 to 7 years of their lives,[54] most mammals stop producing lactase much earlier. Cattle can be weaned from their mothers' milk at 6 months to a year of age.[56] Lambs are regularly weaned around 16 weeks old.[57] Such examples suggest that lactase persistence is a uniquely human phenomenon.
Confounding factors
Some examples exist of factors that can cause the lactase persistence phenotype in the absence of any genetic variant associated with LP. Individuals may lack the alleles for lactase persistence, but still tolerate dairy products in which lactose is broken down by the fermentation process (e.g. cheese, yogurt).[58] Also, healthy colonic gut bacteria may also aid in the breakdown of lactose, allowing those without the genetics for lactase persistence to gain the benefits from milk consumption.[58][59]
References
- 1 2 3 4 5 6 7 8 9 Swallow, D. M. (2003). "Genetics of lactase persistence and lactose intolerance". Annual Review of Genetics. 37: 197–219. doi:10.1146/annurev.genet.37.110801.143820. PMID 14616060.
- 1 2 3 4 5 6 7 Bersaglieri, T.; Sabeti, P. C.; Patterson, N.; Vanderploeg, T.; Schaffner, S. F.; Drake, J. A.; Rhodes, M.; Reich, D. E.; Hirschhorn, J. N. (2004). "Genetic Signatures of Strong Recent Positive Selection at the Lactase Gene". The American Journal of Human Genetics. 74 (6): 1111–1120. doi:10.1086/421051. PMC 1182075. PMID 15114531.
- 1 2 3 4 5 6 Gerbault, Pascale; Liebert, Anke; Itan, Yuval; Powell, Adam; Currat, Mathias; Burger, Joachim; Swallow, Dallas M.; Thomas, Mark G. (2011-03-27). "Evolution of lactase persistence: an example of human niche construction". Philosophical Transactions of the Royal Society B: Biological Sciences. 366 (1566): 863–877. doi:10.1098/rstb.2010.0268. ISSN 0962-8436. PMC 3048992. PMID 21320900.
- ↑ "A worldwide correlation of lactase persistence phenotype and genotypes". BMC Evolutionary Biology. 10. doi:10.1186/1471-2148-10-36.
- 1 2 3 4 5 6 7 8 Heyer, Evelyne; Brazier, Lionel; Ségurel, Laure; Hegay, Tatiana; Austerlitz, Frédéric; Quintana-Murci, Lluis; Georges, Myriam; Pasquet, Patrick; Veuille, Michel (2011-06-01). "Lactase Persistence in Central Asia: Phenotype, Genotype, and Evolution". Human Biology. 83 (3): 379–392. doi:10.3378/027.083.0304. ISSN 0018-7143. PMID 21740154.
- 1 2 Itan, Yuval; Jones, Bryony L.; Ingram, Catherine JE; Swallow, Dallas M.; Thomas, Mark G. (2010-01-01). "A worldwide correlation of lactase persistence phenotype and genotypes". BMC Evolutionary Biology. 10: 36. doi:10.1186/1471-2148-10-36. ISSN 1471-2148. PMC 2834688. PMID 20144208.
- ↑ Durham, William H. (1991). Coevolution: Genes, Culture, and Human Diversity. Stanford, CA: Stanford University Press. ISBN 9780804721561.
- 1 2 3 4 5 Ingram, Catherine J. E.; Mulcare, Charlotte A.; Itan, Yuval; Thomas, Mark G.; Swallow, Dallas M. (2009-01-01). "Lactose digestion and the evolutionary genetics of lactase persistence". Human Genetics. 124 (6): 579–591. doi:10.1007/s00439-008-0593-6. ISSN 0340-6717.
- ↑ Bayoumi, R. a. L.; Flatz, S. D.; Kühnau, W.; Flatz, G. (1982-06-01). "Beja And Nilotes: Nomadic pastoralist groups in the Sudan with opposite distributions of the adult lactase phenotypes". American Journal of Physical Anthropology. 58 (2): 173–178. doi:10.1002/ajpa.1330580208. ISSN 1096-8644.
- 1 2 3 4 5 6 7 8 9 10 11 Tishkoff SA, Reed FA, Ranciaro A, Voight BF, Babbitt CC, Silverman JS, Powell K, Mortensen HM, Hirbo JB, Osman M, Ibrahim M, Omar SA, Lema G, Nyambo TB, Ghori J, Bumpstead S, Pritchard JK, Wray GA, Deloukas P (January 2007). "Convergent adaptation of human lactase persistence in Africa and Europe". Nat. Genet. 39 (1): 31–40. doi:10.1038/ng1946. PMC 2672153. PMID 17159977.
- 1 2 3 4 5 6 7 8 Troelsen JT (May 2005). "Adult-type hypolactasia and regulation of lactase expression". Biochim. Biophys. Acta. 1723 (1–3): 19–32. doi:10.1016/j.bbagen.2005.02.003. PMID 15777735.
- 1 2 3 Wang Y, Harvey CB, Hollox EJ, Phillips AD, Poulter M, Clay P, Walker-Smith JA, Swallow DM (June 1998). "The genetically programmed down-regulation of lactase in children". Gastroenterology. 114 (6): 1230–6. doi:10.1016/S0016-5085(98)70429-9. PMID 9609760.
- ↑ Harvey CB, Wang Y, Hughes LA, Swallow DM, Thurrell WP, Sams VR, Barton R, Lanzon-Miller S, Sarner M (January 1995). "Studies on the expression of intestinal lactase in different individuals". Gut. 36 (1): 28–33. doi:10.1136/gut.36.1.28. PMC 1382348. PMID 7890232.
- ↑ Lloyd M, Mevissen G, Fischer M, Olsen W, Goodspeed D, Genini M, Boll W, Semenza G, Mantei N (February 1992). "Regulation of intestinal lactase in adult hypolactasia". J. Clin. Invest. 89 (2): 524–9. doi:10.1172/JCI115616. PMC 442883. PMID 1737843.
- 1 2 3 4 5 Lokki, A. Inkeri; Järvelä, Irma; Israelsson, Elisabeth; Maiga, Bakary; Troye-Blomberg, Marita; Dolo, Amagana; Doumbo, Ogobara K.; Meri, Seppo; Holmberg, Ville (2011-01-01). "Lactase persistence genotypes and malaria susceptibility in Fulani of Mali". Malaria Journal. 10: 9. doi:10.1186/1475-2875-10-9. ISSN 1475-2875. PMC 3031279. PMID 21235777.
- ↑ Lewinsky, Rikke H.; Jensen, Tine G. K.; Møller, Jette; Stensballe, Allan; Olsen, Jørgen; Troelsen, Jesper T. (2005-12-15). "T −13910 DNA variant associated with lactase persistence interacts with Oct-1 and stimulates lactase promoter activity in vitro". Human Molecular Genetics. 14 (24): 3945–3953. doi:10.1093/hmg/ddi418. ISSN 0964-6906.
- ↑ Jensen, Tine G. K.; Liebert, Anke; Lewinsky, Rikke; Swallow, Dallas M.; Olsen, Jørgen; Troelsen, Jesper T. (2011-10-01). "The −14010*C variant associated with lactase persistence is located between an Oct-1 and HNF1α binding site and increases lactase promoter activity". Human Genetics. 130 (4): 483–493. doi:10.1007/s00439-011-0966-0. ISSN 0340-6717.
- 1 2 3 4 Enattah N. S.; Jensen T. G. K.; Nielsen M.; Lewinski R.; Kuokkanen M.; Rasinpera H.; El-Shanti H.; Kee Seo J.; Alifrangis M.; et al. (2008). "Independent Introduction of Two Lactase-Persistence Alleles into Human Populations Reflects Different History of Adaptation to Milk Culture". American Journal of Human Genetics. 82 (1): 57–72. doi:10.1016/j.ajhg.2007.09.012. PMC 2253962. PMID 18179885.
- ↑ Enattah NS, Sahi T, Savilahti E, Terwilliger JD, Peltonen L, Järvelä I (February 2002). "Identification of a variant associated with adult-type hypolactasia". Nature Genetics. 30 (2): 233–7. doi:10.1038/ng826. PMID 11788828.
- 1 2 3 Ingram, Catherine J. E.; Elamin, Mohamed F.; Mulcare, Charlotte A.; Weale, Michael E.; Tarekegn, Ayele; Raga, Tamiru Oljira; Bekele, Endashaw; Elamin, Farouk M.; Thomas, Mark G. (2007-02-01). "A novel polymorphism associated with lactose tolerance in Africa: multiple causes for lactase persistence?". Human Genetics. 120 (6): 779–788. doi:10.1007/s00439-006-0291-1. ISSN 0340-6717.
- ↑ Imtiaz, F.; Savilahti, E.; Sarnesto, A.; Trabzuni, D.; Al-Kahtani, K.; Kagevi, I.; Rashed, M. S.; Meyer, B. F.; Järvelä, I. (2007-10-01). "The T/G−13915 variant upstream of the lactase gene (LCT) is the founder allele of lactase persistence in an urban Saudi population". Journal of Medical Genetics. 44 (10): e89–e89. doi:10.1136/jmg.2007.051631. ISSN 0022-2593. PMC 2597971. PMID 17911653.
- ↑ Torniainen, Suvi; Parker, M. Iqbal; Holmberg, Ville; Lahtela, Elisa; Dandara, Collet; Jarvela, Irma (2009-01-01). "Screening of variants for lactase persistence/non-persistence in populations from South Africa and Ghana". BMC Genetics. 10: 31. doi:10.1186/1471-2156-10-31. ISSN 1471-2156. PMC 2713916. PMID 19575818.
- 1 2 Gerbault, Pascale; Moret, Céline; Currat, Mathias; Sanchez-Mazas, Alicia; O'Rourke, Dennis; Volm, M; Lorenz, WJ (24 July 2009). "Impact of Selection and Demography on the Diffusion of Lactase Persistence". PLoS ONE. 4 (7): e6369. doi:10.1371/journal.pone.0006369. PMC 2711333. PMID 19629189.
- ↑ Ingram, Catherine J. E.; Raga, Tamiru Oljira; Tarekegn, Ayele; Browning, Sarah L.; Elamin, Mohamed F.; Bekele, Endashaw; Thomas, Mark G.; Weale, Michael E.; Bradman, Neil (2009-12-01). "Multiple Rare Variants as a Cause of a Common Phenotype: Several Different Lactase Persistence Associated Alleles in a Single Ethnic Group". Journal of Molecular Evolution. 69 (6): 579. doi:10.1007/s00239-009-9301-y. ISSN 0022-2844.
- 1 2 3 Enattah, N. S.; Kozlov, A.; Sajantila, A.; Jarvela, I.; Shaat, N.; Groop, L.; et al. (2007). "Evidence of still-ongoing convergence evolution of the lactase persistence T-13910 alleles in humans". The American Journal of Human Genetics. 81 (3): 615–625. doi:10.1086/520705. PMC 1950831. PMID 17701907.
- 1 2 3 Macholdt, Enrico; Lede, Vera; Barbieri, Chiara; Mpoloka, Sununguko W.; Chen, Hua; Slatkin, Montgomery; Pakendorf, Brigitte; Stoneking, Mark (2014-04-14). "Tracing pastoralist migrations to southern Africa with lactase persistence alleles". Current Biology. 24 (8): 875–879. doi:10.1016/j.cub.2014.03.027. ISSN 1879-0445. PMC 5102062. PMID 24704073.
- ↑ Ranciaro, Alessia; Campbell, Michael C.; Hirbo, Jibril B.; Ko, Wen-Ya; Froment, Alain; Anagnostou, Paolo; Kotze, Maritha J.; Ibrahim, Muntaser; Nyambo, Thomas (2014-04-03). "Genetic origins of lactase persistence and the spread of pastoralism in Africa". American Journal of Human Genetics. 94 (4): 496–510. doi:10.1016/j.ajhg.2014.02.009. ISSN 1537-6605. PMC 3980415. PMID 24630847.
- ↑ Itan Y.; Jones B. L.; Ingram C. J. E.; Swallow D. M.; Thomas M. G. (2010). "A worldwide correlation of lactase persistence phenotype and genotypes". BMC Evolutionary Biology. 10 (1): 36. doi:10.1186/1471-2148-10-36. PMC 2834688. PMID 20144208.
- ↑ Troelsen JT, Olsen J, Møller J, Sjöström H (December 2003). "An upstream polymorphism associated with lactase persistence has increased enhancer activity". Gastroenterology. 125 (6): 1686–94. doi:10.1053/j.gastro.2003.09.031. PMID 14724821.
- ↑ Itan, Yuval; Powell, Adam; Beaumont, Mark A.; Burger, Joachim; Thomas, Mark G. (2009). "The Origins of Lactase Persistence in Europe". PLoS Computational Biology. 5 (8): e1000491.
- ↑ Poulter, M.; Hollox, E.; Harvey, C. B.; Mulcare, C.; Peuhkuri, K.; Kajander, K.; Sarner, M.; Korpela, R.; Swallow, D. M. (2003-07-01). "The causal element for the lactase persistence/non-persistence polymorphism is located in a 1 Mb region of linkage disequilibrium in Europeans". Annals of Human Genetics. 67 (Pt 4): 298–311. doi:10.1046/j.1469-1809.2003.00048.x. ISSN 0003-4800. PMID 12914565.
- 1 2 Burger, J.; Kirchner, M.; Bramanti, B.; Haak, W.; Thomas, M. G. (2007). "Absence of the lactase-persistence-associated allele in early Neolithic Europeans". Proceedings of the National Academy of Sciences. 104: 3736–3741. doi:10.1073/pnas.0607187104. PMC 1820653. PMID 17360422.
- ↑ Gerbault, P.; Liebert, A.; Itan, Y.; Powell, A.; Currat, M.; Burger, J.; et al. (2011). "Evolution of lactase persistence: an example of human niche construction". Philosophical Transactions of the Royal Society B: Biological Sciences. 366 (1566): 863–877. doi:10.1098/rstb.2010.0268. PMC 3048992. PMID 21320900.
- 1 2 3 Itan Y, Powell A, Beaumont MA, Burger J, Thomas MG (August 2009). Tanaka MM, ed. "The origins of lactase persistence in Europe". PLoS Comput. Biol. 5 (8): e1000491. doi:10.1371/journal.pcbi.1000491. PMC 2722739. PMID 19714206.
- 1 2 3 Smith GD, Lawlor DA, Timpson NJ, Baban J, Kiessling M, Day IN, Ebrahim S (March 2009). "Lactase persistence-related genetic variant: population substructure and health outcomes". Eur. J. Hum. Genet. 17 (3): 357–67. doi:10.1038/ejhg.2008.156. PMC 2986166. PMID 18797476.
- ↑ Gerbault, Pascale; Moret, Céline; Currat, Mathias; Sanchez-Mazas, Alicia (2009). "Impact of Selection and Demography on the Diffusion of Lactase Persistence". PLoS One. 4 (7): e6369. PMC 2711333.
- ↑ Cook, G. C.; al-Torki, M. T. (1975-07-19). "High intestinal lactase concentrations in adult Arbs in Saudi Arabia". British Medical Journal. 3 (5976): 135–136. doi:10.1136/bmj.3.5976.135. ISSN 0007-1447. PMC 1674005. PMID 1170003.
- ↑ Ingram, Catherine J.; Mulcare, Charlotte A.; Itan, Yuval; Thomas, Marc G.; Swallow, Dallas M. (2009). "Lactase digestion and the evolutionary genetics of lactase persistence". Human Genetics. 124 (6): 587.
- ↑ Bayoumi, R.A.L.; Flatz, S.D.; Kuhnau, W.; Flatz, G. (1982). "Beja and Nilotes: nomadic pastoralist groups in the Sudan with opposite distributions of the adult lactase phenotypes". American Journal of Physical Anthropology. 58 (2): 175. PMID 6810704.
- ↑ Malmstrom H.; Linderholm A.; Liden K.; Stora J.; Molnar P.; Holmlund G.; Jakkobson M.; Gotherstrom A. (2010). "High frequency of lactose intolerance in a prehistoric hunter-gatherer population in northern Europe". BMC Evolutionary Biology. 10: 89. doi:10.1186/1471-2148-10-89. PMC 2862036. PMID 20353605.
- ↑ Swaminathan, N. 2007. Not Milk? Neolithic Europeans Couldn't Stomach the Stuff. Scientific American.
- ↑ Coelho M.; Luiselli D.; Bertorelle G.; Lopes A. I.; Seixas S.; Destro-Bisol G.; Rocha J. (2002). "Microsatellite variation and evolution of human lactase persistence". Human Genetics. 117 (4): 329–339. doi:10.1007/s00439-005-1322-z. PMID 15928901.
- ↑ Aoki K (2001). "Theoretical and Empirical Aspects of Gene–Culture Coevolution". Theoretical Population Biology. 59 (4): 253–261. doi:10.1006/tpbi.2001.1518. PMID 11560446.
- ↑ Olds, Lynne C.; Ahn, Jong Kun; Sibley, Eric (2011-01-01). "−13915*G DNA polymorphism associated with lactase persistence in Africa interacts with Oct-1". Human Genetics. 129 (1): 111–113. doi:10.1007/s00439-010-0898-0. ISSN 0340-6717. PMC 3044188. PMID 20960210.
- ↑ Enattah, N.; et al. (2007). "Evidence of Still-Ongoing Convergence Evolution of the Lactase Persistence T-13910 Alleles in Humans". The American Journal of Human Genetics. 81 (3): 615–25. doi:10.1086/520705. PMC 1950831. PMID 17701907.
- ↑ Myles S.; Bouzekri N.; Haverfield E.; Cherkaoui M.; Dugoujon J. M.; Ward R. (2005). "Genetic evidence in support of a shared Eurasian-North African dairying origin". Human Genetics. 117 (1): 34–42. doi:10.1007/s00439-005-1266-3. PMID 15806398.
- ↑ Mathieson, Iain; et al. (2015). "Genome-wide patterns of selection in 230 ancient Eurasians". Nature. 528 (7583): 501.
- ↑ (Outram et al. 2009
- 1 2 3 4 Anke Liebert (2014). "Evolutionary and molecular genetics of regulatory alleles responsible for lactase persistence" (PDF). University College London. pp. 215–223. Retrieved 11 September 2017.
- 1 2 3 Breton, Gwenna; Schlebusch, Carina M.; Lombard, Marlize; Sjödin, Per; Soodyall, Himla; Jakobsson, Mattias (2014-04-14). "Lactase persistence alleles reveal partial East African ancestry of southern African Khoe pastoralists". Current Biology. 24 (8): 852–858. doi:10.1016/j.cub.2014.02.041. ISSN 1879-0445. PMID 24704072.
- ↑ Hassan, Hisham Y.; et al. (2016). "Genetic diversity of lactase persistence in East African populations". BMC Research Notes. 9 (1): 8. doi:10.1186/s13104-015-1833-1. Retrieved 19 September 2017.
- ↑ Schlebusch, Carina M.; Skoglund, Pontus; Sjödin, Per; Gattepaille, Lucie M.; Hernandez, Dena; Jay, Flora; Li, Sen; Jongh, Michael De; Singleton, Andrew (2012-10-19). "Genomic Variation in Seven Khoe-San Groups Reveals Adaptation and Complex African History" (PDF). Science. 338 (6105): 374–379. doi:10.1126/science.1227721. ISSN 0036-8075. PMID 22997136.
- ↑ Gifford-Gonzalez, Diane (2013). "Animal Genetics and African Archaeology: Why It Matters". African Archaeological Revue. 30: 1–20. doi:10.1007/s10437-013-9130-7. Retrieved 12 September 2017.
- 1 2 Kretchmer N (October 1989). "Expression of lactase during development". Am. J. Hum. Genet. 45 (4): 487–8. PMC 1683494. PMID 2518796.
- ↑ Murray RD, Ailabouni AH, Powers PA, McClung HJ, Li BU, Heitlinger LA, Sloan HR (July 1991). "Absorption of lactose from colon of newborn piglet". Am. J. Physiol. 261 (1 Pt 1): G1–8. PMID 1907103.
- ↑ Menéndez-Buxadera A, Carleos C, Baro JA, Villa A, Cañón J (February 2008). "Multi-trait and random regression approaches for addressing the wide range of weaning ages in Asturiana de los Valles beef cattle for genetic parameter estimation". J. Anim. Sci. 86 (2): 278–86. doi:10.2527/jas.2007-0252. PMID 17998432.
- ↑ Bickell S, Poindron P, Nowak R, Chadwick A, Ferguson D, Blache D (November 2009). "Genotype rather than non-genetic behavioural transmission determines the temperament of Merino lambs". Animal Welfare. 18 (4): 459–466.
- 1 2 Silanikove N, Leitner G, Merin U (2015). "The Interrelationships between Lactose Intolerance and the Modern Dairy Industry: Global Perspectives in Evolutional and Historical Backgrounds". Nutrients (Review). 7 (9): 7312–31. doi:10.3390/nu7095340. PMC 4586535. PMID 26404364.
- ↑ Szilagyi A (2015). "Adaptation to Lactose in Lactase Non Persistent People: Effects on Intolerance and the Relationship between Dairy Food Consumption and Evalution [sic] of Diseases". Nutrients (Review). 7 (8): 6751–79. doi:10.3390/nu7085309. PMC 4555148. PMID 26287234.