Dickinsonia
| Dickinsonia | |
|---|---|
 | |
| Cast of Dickinsonia costata from Australia | |
| Scientific classification | |
| Kingdom: | Animalia |
| Subkingdom: | Eumetazoa |
| Clade: | Bilateria |
| Family: | †Dickinsoniidae |
| Genus: | †Dickinsonia Sprigg, 1947 |
| Species | |
|
See text | |
| Synonyms | |
Dickinsonia is a marine genus of fossils of the Ediacaran biota. The individual Dickinsonia typically resembles a bilaterally symmetrical ribbed oval. Its affinities are presently unknown; its mode of growth is consistent with a bilaterian affinity,[3] though some have suggested that it belongs to the fungi, or even an "extinct kingdom".[4][5][6] The discovery of cholesterol molecules, a signature of animal cells, is evidence that Dickinsonia were animals, and at 571 million to 541 million years ago, may have been among the earliest animals on the planet.[7]
Description

Dickinsonia fossils are known only in the form of imprints and casts in sandstone beds. The specimens found range from a few millimetres to about 1.4 metres (4 ft 7 in) in length, and from a fraction of a millimetre to a few millimetres thick.[8]
They are nearly bilaterally symmetric, segmented, round or oval in outline, slightly expanded to one end (i.e. egg-shaped outline). The rib-like segments are radially inclined towards the wide and narrow ends, and the width and length of the segments increases towards the wide end of the fossil. The segments are separated by thin ridge or groove along the axis of symmetry into right and left halves. But left and right halves are organized in an alternating pattern according to glide reflection symmetry rather than bilateral symmetry;[2][9] thus, these "segments" are isomers.[2][10] This glide reflection is also found in Spriggina, another mysterious organism from approximately the same era.
The segments of Dickinsonia have been described by Adolf Seilacher as self-organising "pneu structures", chambers filled with a liquid at higher than ambient pressure, analogous to a quilted air mattress.[11][12]

Some spectacular fossils attributed to Dickinsonia appear to preserve internal anatomy, believed to represent a tract that both digested food and distributed it throughout the organism.[13]
Similar fossils include Yorgia, which differs in having one end which is segment-free, variously interpreted as a head or hold fast.[14] Another similar fossil is Rutgersella, in which the central groove does not extend to both ends as in Dickinsonia.
The discovery of cholesterol molecules, a signature of animal cells, in Dickinsonia fossils from northwestern Russia, adds to the evidence suggesting Dickinsonia were animals.[7]
Species
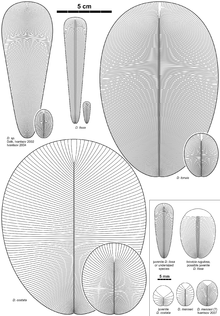
Since 1947, a total of nine species have been described:
- D. costata Sprigg, 1947 (type species)[15]
- D. minima Sprigg, 1949[16]
- D. spriggi Harrington et Moore, 1955[17]
- D. elongata Glaessner et Wade, 1966[18]
- D. tenuis Glaessner et Wade, 1966[18]
- D. lissa Wade, 1972[19]
- D. brachina Wade, 1972[19]
- D. menneri Keller, 1976 =Vendomia menneri Keller, 1976,[20] redefined as Dickinsonia by Ivantsov, 2007[2]
- D. rex Jenkins, 1992[21]
From these, it is possible to consider only four or five of these nine as valid species:
D. costata. (junior synonyms D. minima, D. spriggi, and D. elongata; holotype). Unlike other species, D. costata has comparatively fewer, wider segments/isomers.
D. tenuis (junior synonym D. brachina). Strongly resembles D. costata, but differs from it by more narrow and numerous segments, sparingly lengthened oval form of the body.
D. lissais extremely elongated (up to 15 cm), almost ribbon-like in shape, with numerous thin isomers. The isomers of the head area are short in comparison with those of the rest of the body. The fossil bears a distinct axial ledge consisting of two parallel bands extending from the head region to the posterior end of the body.
D. menneri (junior synonym Vendomia menneri) is a small organism up to 8 mm in length, and strongly resembles juvenile specimens of D. costata with its small number of isomers and well-marked head. D. menneri differs from juvenile D. costata by its slightly more elongated form.
D. rex was erected for selected paratypes of D. elongata. This species is represented by only several very big specimens (up to more than 1 m in length), and does not have a distinct determination. A large size is the major reason for D. rex's species status. Individuals identified as D. rex may simply be large specimens of D. costata and or D. tenuis.
Discovery and naming
The first species and specimens of this fossil organism was first discovered in the Ediacara Member of the Rawnsley Quartzite, Flinders Ranges in South Australia. Additional specimens of Dickinsonia are also known from the Mogilev Formation in the Dniester River Basin of Podolia, Ukraine[22] and from the Lyamtsa, Verkhovka, Zimnegory and Yorga Formations in the White Sea area of the Arkhangelsk Region, and Chernokamen Fm. of the Central Urals, Russia.[8] These deposits have been dated to 558–555 Myr.[23]
Reg Sprigg, the original discoverer of the Ediacaran biota in Australia,[15] described Dickinsonia, naming it after Ben Dickinson, then Director of Mines for South Australia, and head of the government department that employed Sprigg.
Body fossils
As a rule, Dickinsonia fossils are preserved as negative impressions on the bases of sandstone beds. Such fossils are imprints of the upper sides of the benthic organisms that have been buried under the sand.[24][25] The imprints formed as a result of cementation of the sand before complete decomposition of the body. The mechanism of cementation is not quite clear; among many possibilities, the process could have arisen from conditions which gave rise to pyrite "death masks"[25] on the decaying body, or perhaps it was due to the carbonate cementation of the sand.[26] The imprints of the bodies of organisms are often strongly compressed, distorted, and sometimes partly extend into the overlying rock. These deformations appear to show attempts by the organisms to escape from the falling sediment.[10][27][28]
Rarely, Dickinsonia have been preserved as a cast in massive sandstone lenses, where it occurs together with Pteridinium, Rangea and some others.[23][20][29][30] These specimens are products of events where organisms were first stripped from the sea-floor, transported and deposited within sand flow.[23][30] In such cases, stretched and ripped Dickinsonia occur. The first such specimen was described as a separate genus and species, Chondroplon bilobatum[31] and later redefined as Dickinsonia.[1]
Trace fossils
Several trace fossils, including Epibaion and Phylozoon, have been interpreted as feeding impressions of Dickinsonia and its relatives. Such fossils consist of large, rounded impressions of the undersides of these animals.[10][27][32] They may be impressions the organism made while it rested on the sediment surface – perhaps by secreting slime in order to form a platform on the underlying microbial mat,[33] or by sitting and dissolving the underlying microbes in order to devour them.[33][34] They have also been interpreted as "tumble tracks" created by an organism rolling along the sea floor, perhaps as it was buffeted by currents,[33] and as the bases of lichens or "mushrooms arranged in fairy rings".[33] However, in some cases these trackway imprints overlap. Ridges apparently produced by the channelling of sediment in digestive tubes seem to indicate that the trackways do indeed represent feeding traces; the sedimentary disturbance expected of tumbling-induced impressions is not observed.[10][27]
Classification
The affinities of Dickinsonia are uncertain. It has been variously interpreted as a jellyfish, coral, polychaete worm, turbellarian, mushroom, xenophyophoran protist, sea anemone, lichen,[33][35] and even a close ancestor of the chordates.[36] Genera such as Rutgersella, Yorgia and Marywadea somewhat resemble Dickinsonia, and may be related. However, it is possible that Dickinsonia falls into a group of organisms that became extinct before the Cambrian. Its construction is loosely similar to other Ediacaran organisms, and the similarity of their architecture suggests that dickinsoniamorphs may belong in a clade with Charnia and other rangeomorphs.[37] Paleontologist Adolph Seilacher even went so far as to suggest that most of the Ediacaran fauna represents a separate Kingdom termed "Vendozoa" (now: "Vendobionta") that thrived just before most of the modern multicellular animal phyla appeared in the fossil record.[5]
There is a strong argument that the organism is more derived than a sponge, but less so than a eumetazoan. The organism could clearly move, evidenced by its association with trackways which could only have been produced by feeding.[38] However, it lacks any convincing evidence for a mouth, anus or gut, and appears to have fed by absorption on its bottom surface. The placozoans are simple animals which feed with their soles and are phylogenetically between sponges and eumetazoa; this suggests that Dickinsonia may have been a stem-group placozoan, or somewhere more crownwards than sponges on the eumetazoan stem. [38][39]
Greg Retallack proposed that some Ediacaran fossils were lichens based on their unusual resistance to post-burial compaction,[33][40] He proposes that the decay mode of the organisms is most similar to that of leaves, fungi or lichens, unlike soft-bodied animals which clot and distort as they wilt and decay.[33] A detailed study of paleosols with Dickinsonia preserved in life position suggests, according to Retallack, that Dickinsonia could have lived on dry land.[41] The proposal is not widely accepted in the scientific community.[42][43] The September 2018 paper announcing the discovery of cholesteroids in Dicksonia fossils states: "Lichen-forming fungi only produce ergosteroids, and even in those that host symbiotic algae, ergosteroids remain the major sterols. Dickinsonia contained no or a maximum of only 0.23% ergosteroids, conclusively refuting the lichen hypothesis".[7]
In 2017 it was determined that Dickinsonia is an Eumetazoan but a more accurate affiliation was not established.[44] In a subsequent study Hoyal Cuthill & Han (2018) assigned Dickinsonia to the extinct animal group Petalonamae (placed as sister group to the Eumetazoa), which also included the genera Stromatoveris, Arborea and Pambikalbae, as well as rangeomorphs and erniettomorphs.[45]
Paleobiology

Dickinsonia displayed isometric, indeterminate growth – that is to say, they kept growing until they were covered with sediment or otherwise killed.[33] They spent most, if not all, of their lives with most of their bodies firmly anchored to the sediment, although they may have moved from resting-place to resting-place.[38] Their mode of anchorage may have been oyster-like concretion, lichen-like rooting with rhizines, or fungus-like attachment to an underground network of hyphae.[33]
The organisms are preserved in such a way that their resistant parts must have been a sturdy biopolymer (such as keratin) rather than a brittle mineral (such as calcite or a pyritized death mask).
Halo-like "reaction rims" surround specimens;[33] adjacent specimens deform as if to avoid entering their neighbour's halo, suggesting they competed with one another.[33] No body fossils have been found to overlap.[33]
See also
References
- 1 2 Hofmann, Hans J. (1988). "An alternative interpretation of the Ediacaran (Precambrian) chondrophore Chondroplon Wade". Alcheringa. 12 (4): 315–318. doi:10.1080/03115518808619130.
- 1 2 3 4 Ivantsov, A. Yu (2007). "Small Vendian transversely Articulated fossils". Paleontological Journal. 41 (2): 113–122. doi:10.1134/S0031030107020013.
- ↑ Gold, D. A.; Runnegar, B.; Gehling, J. G.; Jacobs, D. K. (2015). "Ancestral state reconstruction of ontogeny supports a bilaterian affinity for Dickinsonia". Evolution & Development. 17 (6): 315–397. doi:10.1111/ede.12168.
- ↑ Pflug (1973). "Zur fauna der Nama-Schichten in Südwest-Afrika. IV. Mikroscopische anatomie der petalo-organisme". Paleontographica (B144): 166–202.
- 1 2 Seilacher, Adolf (1992). "Vendobionta and Psammocorallia: lost constructions of Precambrian evolution". Journal of the Geological Society, London. 149 (4): 607–613. doi:10.1144/gsjgs.149.4.0607. Retrieved 2007-06-21.
- ↑ McMenamin M. (1998). The Garden of Ediacara. New York: Columbia University Press. ISBN 978-0-231-10559-0. OCLC 228271905. It lived during the late Ediacaran (part of Precambrian). Dinosaurs a visual encyclopedia. New York. ISBN 9781465469489.
- 1 2 3 Bobrovskiy, Ilya; Hope, Janet M.; Ivantsov, Andrey; Nettersheim, Benjamin J.; Hallmann, Christian; Brocks, Jochen J. (20 September 2018). "Ancient steroids establish the Ediacaran fossil Dickinsonia as one of the earliest animals". Science. 361 (6408): 1246–1249. doi:10.1126/science.aat7228.
- 1 2 Fedonkin M. A.; Gehling J. G.; Grey K.; Narbonne G. M.; Vickers-Rich P. (2007). The Rise of Animals. Evolution and Diversification of the Kingdom Animalia. Johns Hopkins University Press. p. 326. ISBN 978-0-8018-8679-9.
- ↑ Ivantsov, A. Yu (2012). "Becoming metamery and bilateral symmetry in Metazoa: way of Proarticulata". Morphogenesis in the individual and historical development: symmetry and asymmetry.: 16–17.
- 1 2 3 4 Ivantsov, A. Y. (2011). "Feeding traces of Proarticulata — the Vendian metazoa". Paleontological Journal. 45 (3): 237–248. doi:10.1134/S0031030111030063.
- ↑ Seilacher, Adolf (1991). Schmidt-Kittler, Norbert; Vogel, Klaus, eds. Self-Organizing Mechanisms in Morphogenesis and Evolution. Constructional Morphology and Evolution. Springer. pp. 251–271. ISBN 978-3-642-76158-4.
- ↑ Seilacher, Adolf (July 1989). "Vendozoa: Organismic construction in the Proterozoic biosphere". Lethaia. 22: 229–239. doi:10.1111/j.1502-3931.1989.tb01332.x.
- ↑ Ivantsov, A. Y. (2004). "New Proarticulata from the Vendian of the Arkhangel'sk Region" (PDF). Paleontological Journal. 38 (3): 247–253. Archived from the original (PDF) on 2007-09-27. Retrieved 2007-09-27.
- ↑ Seilacher, A. (2013). "Patterns of macorevolution through the Phanerozoic". Palaeontology. 56: 1273–1286.
- 1 2 Sprigg, Reg C. (1947). "Early Cambrian (?) Jellyfishes from the Flinders Ranges, South Australia" (PDF). Trans. Roy. Soc. S. Aust. 71: 212–24. Archived from the original (PDF) on 2007-09-29.
- ↑ Sprigg, R.C. (1949). "Early Cambrian "Jellyfishes" of Ediacara, South Australia and Mount John, Kimberley District, Western Australia" (PDF). Transactions of the Royal Society of South Australia. 73: 72–99.
- ↑ Harrington, N. J.; Moore. R. C. (1955). "Kansas Pennsylvanian and other jellyfishes". Bull. Kansas geol. Surv. 114 (5): 153–163.
- 1 2 Glaessner, M.F.; Wade, M. (1966). "The late Precambrian fossils from Ediacara, South Australia" (PDF). Palaeontology. 9 (4): 599.
- 1 2 Wade, M. (1972). "Dickinsonia: Polychaete Worms from the Late Precambrian Ediacara Fauna, South Australia". Mem. Queensl. Mus. 16 (2): 171–190.
- 1 2 B. M. Keller; M. A. Fedonkin (1976). "New Records of Fossils in the Valdaian Group of the Precambrian on the Syuz'ma River" (PDF). Izv. Akad. Nauk SSSR, Ser. Geol. (in Russian). 3: 38–44. Archived from the original (PDF) on 2007-09-27.
- ↑ Jenkins, R. J. F. (1992). "Functional and ecological aspects of Ediacarian assemblages". In Lipps, J., and Signor, P. W. Origin and early evolution of the Metazoa. New York: Springer. pp. 131–176. ISBN 0-306-44067-9. OCLC 231467647.
- ↑ Fedonkin, M. A. (1983). "Non-skeletal fauna of Podoloia, Dniester River valley". In Velikanov, V. A.; Assejeva, E. A.; Fedonkin, M. A. The Vendian of the Ukraine (in Russian). Kiev: Naukova Dumka. pp. 128–139.
- 1 2 3 Grazhdankin, Dima (2004). "Patterns of distribution in the Ediacaran biotas: facies versus biogeography and evolution" (PDF). Paleobiology. 30 (2): 203–221. doi:10.1666/0094-8373(2004)030<0203:PODITE>2.0.CO;2.
- ↑ Ivantsov, A. Y. (2009). "A New Reconstruction of Kimberella, a Problematic Vendian Metazoan". Paleontological Journal. 43 (6): 601–611. doi:10.1134/S003103010906001X.
- 1 2 Gehling, J.G. (1999). "Microbial mats in terminal Proterozoic siliciclastics; Ediacaran death masks". PALAIOS. 14 (1): 40–57. doi:10.2307/3515360. JSTOR 3515360.
- ↑ Serezhnikova, E.A. (2011). "Microbial Binding as a Probable Cause of Taphonomic Variability of Vendian Fossils: Carbonate Casting?". Lecture Notes in Earth Sciences. 131: 525–535. doi:10.1007/978-3-642-10415-2_31. ISBN 978-3-642-10414-5.
- 1 2 3 Ivantsov, A.Yu. (2013). "Trace Fossils of Precambrian Metazoans "Vendobionta" and "Mollusks"". Stratigraphy and Geological Correlation. 21 (3): 252–264. doi:10.1134/S0869593813030039.
- ↑ Runnegar, Bruce (1982). "Oxygen requirements, biology and phylogenetic significance of the late Precambrian worm Dickinsonia, and the evolution of the burrowing habit". Alcheringa: an Australasian Journal of Palaeontology. 6 (3): 223–239. doi:10.1080/03115518208565415.
- ↑ Keller, B. M.; Fedonkin, M. A. (1977). "New organic fossil finds in the Precambrian Valday series along the Syuz'ma River". International Geology Review. 19 (8): 924–930. doi:10.1080/00206817709471091.
- 1 2 Gehling, J.G.; Droser, M.L. (2013). "How well do fossil assemblages of the Ediacara Biota tell time?". Geology. 41 (4): 447–450. doi:10.1130/G33881.1.
- ↑ Wade, M. (1971). "Bilateral Precambrian Chondrophores from the Ediacara Fauna, South Australia". Proceedings of the Royal Society of Victoria. 84 (1): 183–188.
- ↑ Ivantsov, A.Y.; Malakhovskaya, Y.E. (2002). "Giant Traces of Vendian Animals" (PDF). Doklady Earth Sciences. 385 (6): 618–622. Archived from the original (PDF) on 2007-07-04. Retrieved 2008-02-24.
- 1 2 3 4 5 6 7 8 9 10 11 12 Retallack, G.J. (2007). "Growth, decay and burial compaction of Dickinsonia, an iconic Ediacaran fossil" (PDF). Alcheringa: an Australasian Journal of Palaeontology. 31 (3): 215–240. doi:10.1080/03115510701484705.
- ↑ Jensen, Sören; Droser, Mary L.; Gehling, James G. (2 May 2005). "Trace fossil preservation and the early evolution of animals". Palaeogeography, Palaeoclimatology, Palaeoecology. 220 (1–2): 19–29. doi:10.1016/j.palaeo.2003.09.035.
- ↑ Retallack, Gregory J. (2004) "Death, Decay and Destruction of Dickinsonia". Geological Society of America Abstracts with Programs, Vol. 36, No. 5, p. 521
- ↑ Dzik, Jerzy. (2000) "The Origin of the Mineral Skeleton in Chordates." in Max Knobler Hecht, Ross J. MacIntyre and Michael T. Clegg, eds. Evolutionary Biology Vol. 31. Pp. 105-46. Springer. ISBN 0-306-46178-1 – URL retrieved February 10, 2007
- ↑ Brasier, M.; Antcliffe, J. (2 August 2004). "Decoding the Ediacaran Enigma". Science. 305 (5687): 1115–1117. doi:10.1126/science.1102673. PMID 15326344.
- 1 2 3 Sperling, Erik; Vinther, Jakob; Pisani, Davide; Peterson, Kevin (2008). "A placozoan affinity for Dickinsonia and the evolution of Late Precambrian metazoan feeding modes" (PDF). In Cusack, M; Owen, A; Clark, N. Programme with Abstracts. Palaeontological Association Annual Meeting. 52. Glasgow, UK. p. 81.
- ↑ Sperling, Erik; et al. (2008). "A Placozoan Affinity for Dickinsonia and the Evolution of Late Precambrian Metazoan Feeding Modes". Geological Society of America Abstracts with Programs. 40 (6): 508.
- ↑ Retallack, G.J. (1994). "Were the Ediacaran fossils lichens?". Paleobiology. 17: 523–544.
- ↑ Retallack, G.J. (3 January 2013). "Ediacaran life on land". Nature. 493 (7430): 89–92. doi:10.1038/nature11777. PMID 23235827.
- ↑ Waggoner, B.M. (1995). "Ediacaran Lichens: A Critique". Paleobiology. Paleobiology, Vol. 21, No. 3. 21 (3): 393–397. JSTOR 2401174.
- ↑ Waggoner, B.; Collins, A. G. (2004). "Reductio Ad Absurdum: Testing The Evolutionary Relationships Of Ediacaran And Paleozoic Problematic Fossils Using Molecular Divergence Dates". Journal of Paleontology. 78 (1): 51–61. doi:10.1666/0022-3360(2004)078<0051:RAATTE>2.0.CO;2.
- ↑ Hoekzema, Renee S.; Brasier, Martin D.; Dunn, Frances S.; Liu, Alexander G. (2017-09-13). "Quantitative study of developmental biology confirms Dickinsonia as a metazoan". Proc. R. Soc. B. 284 (1862): 20171348. doi:10.1098/rspb.2017.1348. PMID 28904140.
- ↑ Cuthill, Jennifer F. Hoyal; Han, Jian (2018). "Cambrian petalonamid Stromatoveris phylogenetically links Ediacaran biota to later animals". Palaeontology. in press. doi:10.1111/pala.12393.