Koskinonodon
Koskinonodon is an extinct genus of large temnospondyl amphibians. These animals were part of the family called Metoposauridae, which filled the crocodile-like predatory niches in the late Triassic.[2] It may have reached up to 3 m (10 ft) in length, with a 65-cm-long skull.[3] It was an ambush hunter, snapping up anything small enough to fit in its huge jaws.[3] It was very common during the Late Triassic (Norian age) in what is now the American Southwest.[4]
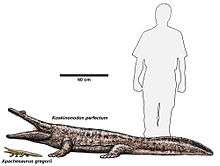
| Koskinonodon | |
|---|---|
 | |
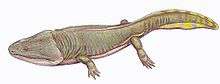
| K. perfecta skeleton at the AMNH | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Order: | †Temnospondyli |
| Suborder: | †Stereospondyli |
| Family: | †Metoposauridae |
| Genus: | †Koskinonodon Branson and Mehl, 1929 |
| Species | |
| |
| Synonyms | |
| |
History of discovery

Known Koskinonodon fossils have been found in the United States, especially the Chinle Formation of Petrified Forest National Park and the Placerias Quarry in Arizona, the Garita Creek Formation of central New Mexico (the quarry at Lamy), the Petrified Forest Member of northern New Mexico, the Bluewater Creek Formation of western New Mexico, the New Oxford Formation of Pennsylvania, the Tecovas Formation of western Texas, and the Popo Agie Formation of the Chugwater Group of Wyoming.[5][6]
Some metoposaurids, like M. diagnosticus and A. Iyazidi, share so many characteristics with K. perfectus and "Metoposaurus" bakeri that it was difficult to differentiate them and create an accurate phylogeny.[7] Many paleontologists think that lachrymal position is a poor taxonomic indicator as it is so difficult to determine in many specimens. However, metoposaurids vary in so few taxonomic characteristics that it is unwise to abandon any that may be important and distinguishing.[7]
The genus was formerly named Buettneria by Case in 1922, but in 2007, B.D. Mueller realized that the name Buettneria had already been given to a bush cricket from the Republic of Congo by Karsch (1889), so he made the genus Koskinonodon the earliest available unpreoccupied name for the temnospondyl.[8] Lucas et al. (2007), however, petitioned the ICZN to suppress Karsch's name in favor of Case's name, citing evidence that the amphibian name is much more well known and widely used (the authors cited 75 uses of the name in scientific literature and books over the last 85 years), and that the bush cricket name had been seldom used in technical literature.[9] However, ICZN Opinion 2255 issued in 2010 rejected the petition.[10]
Gee et al. (2019) redescribed the holotypes of the two nominal Anarchism species and synonymized Anaschisma[11], Buettneria, and Borborophagus with Koskinonodon.[12]
Species
- Koskinonodon perfectus: This species differs from K. bakeri in that the lachrymal enters the orbit.[7] It was named by Case in 1922.
- Koskinonodon bakeri: The lachrymal does not contact the orbit, as it does in K. perfectus. In fact, it evolves to be even father from the orbit than it was in the pair's most recent common ancestor.[7] This is one of the only characteristics that distinguish it from K. perfectus.[13] It was named in 1931 by Case.
Description
The skull of Koskinonodon differs from the skulls of other closely related organisms in a few key ways. It is wider overall and features eye sockets that are very anterior and forward oriented, more so than its contemporaries.[14] Also, the skull has prominent slime canals, which are used for transporting mucus, as well as large external nares.[15] In addition, the upper jaw is relatively weak and thin, used only for holding teeth.[15] Their large jaws could have held many teeth at once, maybe even over 100 on each side of the upper and lower jaws, but the actual number varies constantly over the animal's lifetime due to natural causes such as fighting, eating, disease, etc.[15] The sheer size of the skull is one of the most defining traits for the genus Koskinonodon and its species.[15]
Other more minor skull traits characterize Koskinonodon as well: elongation of the lachrymal, shortening of the prefrontal, reduction of the interclavicle, and the most characteristic is the center lachrymal entering margin of the orbit.[16] While the shortened prefrontal is a characteristic of the family Metoposauridae, it is shorted even more in Koskinonodon.[2] The interclavicle is reduced in the way that it has many hexagonal pits as well as grooves and ridges.[2] The skull of Koskinonodon is also covered in this reticulate ornamentation.[17] Some researchers believe that Koskinonodon has a shorter posterior process of the interclavicle, which may discriminate it from other closely related species, while others believe that there is not enough information to make that distinction.[2] Koskinonodon also has ossified opisthotics, the more posterior of the bones surrounding the inner ear.[17]
Koskinonodon had sharp, pointy teeth for catching and killing prey. They had marginal teeth as well as larger teeth on the palate, specifically palatine and ectopterygoid teeth.[2] They had two basic types: large with shallow grooves and small with deeper grooves. These grooves run lengthwise down the teeth and aided the animals in catching prey.[18] The teeth are elongated labio-lingually at their base and opposite, mediodistally, at their tip. In the middle, they are not elongated either way but instead circular.[18] These dental adaptations enhanced Koskinonodon’s ability to capture prey; the teeth are optimized for piercing prey and not allowing it to escape, resisting the bending force applied by the struggling organism, and propagating cracks in the hard parts of the object, such as bone, allowing for easier eating and digestion.[18]
Koskinonodon had a wide, wedge-shaped, powerful tail to assist it with swimming, hunting and likely defense.[16] It was not long like the crocodiles of today, but more likely short and strong to enable it to quickly spring up from hiding and capture prey before it escapes.[16] The legs of Koskinonodon display a sprawling stance and short legs with 4-digits on the front and 5 on the back limbs.[16] Koskinonodon likely spent a lot of time motionless, waiting for prey, so small, stubby legs are adaptive.[19]
Paleobiology

The hunting style of Koskinonodon involved lying at the bottom of a shallow swamp, waiting for a fish, crustacean, smaller amphibian, or even a young phytosaur to wander by.[3] When it spotted prey, it used its huge jaws to engulf and consume them.[3] A few particular adaptations suggest Koskinonodon had this aquatic lifestyle. First, they had lateral lines formed by the sensory sulci. These are useful for detecting changes in water pressure made by the swimming motions of nearby organisms. Their sprawling limbs were also adapted for water. They would not move quickly or efficiently on land, although they may have done it to find another water pool with more food or other resource. Mass graves have been found, thought to be a result of a group of these animals gathering together in a withering water pool during a drought and all perishing because the water was never replenished.[15]
Paleoecology
They lived mostly in the late Triassic; by the time the Jurassic began, most temnospondyls, Koskinonodon included, were gone.[20] It is likely that they went extinct during the Triassic-Jurassic extinction event, along with the majority of other large amphibians, the class of Conodonts and 34% of all marine genera. It is unknown what caused this mass extinction; theories include huge volcanic eruptions, climate change, oceanic acidification, and even another asteroid impact. It is known, however, that over half of the species living on Earth at that time went extinct from this event.[21] It lived alongside lots of other smaller amphibians, and its fossils are also commonly found with phytosaur fossils.[15] The best conditions for fossilization occur in river valleys or floodplains where deposition is occurring, and this animal likely lived in similar shallow, swampy habitats. As it follows, Koskinonodon is famous for having extremely well preserved fossils, and they are often found in groups.[15]
References
- Bryan M. Gee; William G. Parker; Adam D. Marsh (2019). Redescription of Anaschisma (Temnospondyli: Metoposauridae) from the Late Triassic of Wyoming and the phylogeny of the Metoposauridae. Journal of Systematic Palaeontology in press. doi:10.1080/14772019.2019.1602855.
- Brusatte, Stephen L (2015). "A New Species of Metoposaurus from the Late Triassic of Portugal and Comments on the Systematics and Biogeography of Metoposaurid Temnospondyls". Journal of Vertebrate Paleontology. doi:10.1080/02724634.2014.912988.
- http://palaeos.com/vertebrates/temnospondyli/metoposauroidea.html
- Hunt, A.P. 1989. Comments on the taxonomy of North American metoposaurids and a preliminary phylogenetic analysis of the family Metoposauridae, in Lucas, SG and Hunt, AP, eds., Dawn of the age of dinosaurs in the American Southwest, pp. 292-300 Albuquerque, New Mexico Museum of Natural History.
- Romer, A. S. 1947. Review of the Labyrinthodontia. Bulletin of the Museum of Comparative Zoology, 99, 1–368.
- Branson, E. B. & Mehl, M. G. 1929. Triassic amphibians from the Rocky Mountain region. The University of Missouri Studies, 4, 154–253.
- Lucas, Spencer G; Hunt, Adrian P (1989). Dawn of the Age of Dinosaurs in the American Southwest. 1. New Mexico Museum of Natural History.
- Mueller, B.D. (2007). "Koskinonodon Branson and Mehl, 1929, a replacement name for the preoccupied temnospondyl Buettneria Case, 1922". Journal of Vertebrate Paleontology. 27 (1): 225. doi:10.1671/0272-4634(2007)27[225:KBAMAR]2.0.CO;2.
- Lucas, SG; Rinehart, LF; Spielmann, JA (2008). "Comments on the proposed conservation of Buettneria Case, 1922 (Amphibia) 2 (Case 3420)". Bulletin of Zoological Nomenclature. 65 (3): 218–219.
- http://iczn.org/content/opinion-2255-case-3420-buettneria-case-1922-amphibia-generic-name-not-conserved
- Chowdhury, T. R. 1965. A new metoposaurid amphibian from the Upper Triassic Maleri Formation of Central India. Philosophical Transactions of the Royal Society of London, Series B, 250, 1–52.
- Bryan M. Gee; William G. Parker; Adam D. Marsh (2019). Redescription of Anaschisma (Temnospondyli: Metoposauridae) from the Late Triassic of Wyoming and the phylogeny of the Metoposauridae. Journal of Systematic Palaeontology in press. doi:10.1080/14772019.2019.1602855.
- Parker, W.G.; Martz, J.W. (2010). "The Late Triassic (Norian) Adamanian–Revueltian tetrapod faunal transition in the Chinle Formation of Petrified Forest National Park, Arizona". Earth and Environmental Science Transactions of the Royal Society of Edinburgh: 231–260. doi:10.1017/S1755691011020020.
- Moodie, R. L. 1908. The lateral line system in extinct Amphibia. Journal of Morphology, 19, 511–540.
- Peredo, Carlos. "Koskinonodon Perfectus". Encyclopedia of Life. Missing or empty
|url=(help) - "Palaeos Vertebrates Temnospondyli: Trematosauria: Metoposaurs". Palaeos.com. Missing or empty
|url=(help) - Rinehart, Larry F; Lucas, Spencer G (2013). "THE FUNCTIONAL MORPHOLOGY OF DERMAL BONE ORNAMENTATION IN TEMNOSPONDYL AMPHIBIANS". Museum of Natural History and Science (The Triassic System).
- New Mexico Museum of Natural History & Science. "The Triassic System: New Developments in Stratigraphy and Paleontology". Cite journal requires
|journal=(help) - Branson, E. B. 1905. Structure and relationships of American − Labyrinthodontia. The Journal of Geology, 13, 568–610.
- McHugh, Julia Beth (2012). "TEMNOSPONDYL ONTOGENY AND PHYLOGENY, A WINDOW INTO TERRESTRIAL ECOSYSTEMS DURING THE PERMIAN-TRIASSIC MASS EXTINCTION". Graduate College of the University of Iowa.
- "End-Triassic extinction". Encyclopedia Britannica.
External links
| ||||||||||||||||||||
| ||||||||||||||||||||
| ||||||||||||||||||||
| ||||||||||||||||||||
| ||||||||||||||||||||
| ||||||||||||||||||||
| ||||||||||||||||||||
| ||||||||||||||||||||