Memory and social interactions
Memory underpins and enables social interactions in a variety of ways. In order to engage in successful social interaction, people must be able to remember how they should interact with one another, whom they have interacted with previously, and what occurred during those interactions. Failure in any of these areas can lead to social rejection or violent conflict, which can be detrimental to a person's well being.
Memory for faces
The ability to visually identify previous social partners is essential for successful interactions because it aids in recognizing which partners can and cannot be trusted. In humans, this is accomplished by facial recognition. Research suggests that humans are born with an innate ability to process other human faces. In one study, Pascalis, et al. (1995) found that four-day-old neonates (infants) prefer to look at their mothers' faces rather than at a stranger's.[1] This finding suggests that neonates are able to remember, recognize, and differentiate between faces. Further research suggests that humans prefer to attend to faces rather than non-face alternatives.[2] Such specialized processing for faces aids in the encoding of memory for people. This preference is one explanation for why humans are more proficient at memorizing faces than non-faces.
Brain structures
There are several areas of the brain associated with this highly specialized form of memory.
Fusiform face area
The fusiform face area (FFA) is associated with facial recognition. It is generally located on the fusiform gyrus in the temporal lobe, but its precise location varies between individuals.[3] The FFA has been found to show a degree of lateralization.[3] Typically, it is larger in the right hemisphere of the brain.[3] It is widely accepted that the FFA is involved in the encoding and retrieval of memory for faces and other familiar categorical processing.[4]
Damage to the FFA has been shown to lead to severe deficits in facial recognition and processing.[5] These deficits can lead to difficulty in maintaining normal social relationships over an extended period.
Amygdala
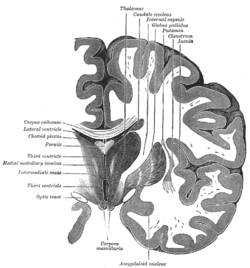
The amygdala is one of the major structures in the limbic system.[6] It is also known to play a role in the processing of and memory for emotional events.[7]
One study, performed by Adolphs, et al. (1998),[8] found striking differences in perceived trustworthiness and approachability ratings between subjects with complete bilateral amygdala damage and a control group. The results showed that all of the amygdally impaired subjects rated unfamiliar faces as much more trustworthy and approachable than familiar faces. Furthermore, the impaired subjects rated faces as trustworthy which the control group considered dubious. These findings illustrate a large difference in the amygdally impaired subjects' perception of faces. The same study went on to determine whether or not the difference generalizes to an auditory description of a person. The impaired subjects made entirely normal perceptions in this task.[8] Overall, the research suggests that the amygdala is important for the making and retrieval of social judgements.
Hippocampus
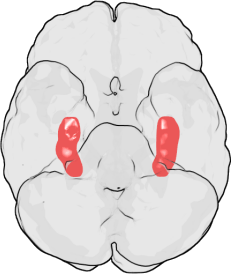
The hippocampus is a structure located in the medial temporal lobe. It is thought to be involved in the encoding, consolidation and retrieval of memories.[9] This structure's function is probably best known from specific case studies, the most famous being Henry Molaison, formerly known as patient H.M., whose medial temporal lobes were bilaterally removed in an attempt to cure his epileptic seizures. As a result, he experienced severe anterograde amnesia.[10]
Difficulties with facial recognition
Several disorders or impairments have been found to disrupt an individual's ability to successfully process faces.
Autism spectrum disorders
Autism spectrum disorders (ASD) are neurological development disorders characterized by repetitive behaviours and impaired social skills.[11] Patients with ASD are also characterized by over-selectivity, which is a tendency to attend to only a few stimuli.[12] Over-selectivity causes memory encoding problems, as relevant information is not attended to, and thus not stored in memory. These encoding problems are associated with an impaired memory for faces,[13] which is in turn associated with impairments in social functioning.[14]

Prosopagnosia
Prosopagnosia is an inability to identify faces and face-like objects. This represents a failure to encode incoming visual information. Neurological studies indicate that prosopagnosia is associated with bilateral lesions of the central visual system, primarily located in the mesial occipitotemporal region.[15] The inability to correctly recognize a face can have detrimental consequences for building social relationships due to the fact that recognition is required for associating feelings or experiences with a stimulus.[14]
Memory for previous interactions
Memory for previous interactions is critical for successful long-term social relationships. This is largely due to a human expectation of reciprocity,[16] which requires humans to have a better memory for favours or debts owed.[16] These memories play an important role in deciding whether or not a human interacts with another in the future, as they contribute to the construction of a person's reputation. Milinski, Semmann & Krambeck (2002) have demonstrated, through the use of public goods games and indirect reciprocity games, that humans are less likely to interact with those who have a reputation for not equivalently returning the favour.[17]
Research indicates that long-term memory can directly influence a person's choice of strategy during interaction.[18] Participants with normal memory functioning were found to use a wider variety of techniques in the prisoner's dilemma task.[18] Thus, memory constraints alter the strategies used during social interactions.[19]
Evolutionary psychologists have shown that memories for social interactions are subject to natural selection.[20][21] As demonstrated through free recall and recognition tasks, humans are able to discern between cooperators and cheaters in social interactions.[22][23]
Impression formation
The impressions humans leave each other with after interactions, whether active or passive, help to build reputations.[17]
A study by Espejo (2003) has demonstrated that the extent of a social interaction does not significantly affect short-term memory retention for that interaction.[24] This indicates that the processes by which impressions are formed do not take much time to occur. Furthermore, as these results were demonstrated in rats with lesioning in the medial prefrontal cortex, a key brain structure for social interaction, they suggest a multi-component system of memory.[24] However, a study by Belmore (1987) on impression formation and serial position indicated that behaviours from early in an acquaintance are no better recalled than later events.[25]
Expectations about a person can influence what is remembered about that individual after an initial interaction. Therefore, such information affects the impression one person makes on another. For example, Srull (1981) found that people had better recall for memories of a person acting in a way opposite to previous expectations.[26] This was attributed to the repeated retrieval of such memories in attempts to reconcile them with the person's reputation.[26][27]
Furthermore, the memories that contribute to an impression appear to be influenced by negativity and the source of behaviour.[28] Research on the subject indicates that people are more likely to remember negative behaviour when it is perceived as coming from a moral or dispositional source, and positive behaviour when seen as coming from either a person's ability or a need to suit the situation.[28][29] Positive behaviours result from a desire to appear socially acceptable, whereas negative behaviours are more revealing of the individual's personality.[28] Thus, negative behaviours lead to more questioning of the individual's actual personality, which in turn results in more methodical processing of that person's behaviour.[25] This is believed to produce better memory for events by forming more associative links to the memory of that behaviour.[26] However, positive behaviour may also be subject to this improved memory if there is reason to doubt the behaviour.[28] If a critical assessment of an individual's actions results in a contrast to their positive image, the amount of interaction with that person will decrease.[28]
Memory for cheaters and cooperators
From a psychological standpoint, friends and foes have been defined in a number of ways, but do not have a consistent operational definition. One definition of friends and foes is as cheaters and non-cheaters, with non-cheaters also referred to as cooperators.[30][31][32] Cheaters are defined as those who benefit from cooperation, but endure no costs, thereby discouraging cooperators. Cheaters benefit from others' cooperation without the costs of reciprocation. Cooperation is risky because those who cooperate gamble with the possibility of being exploited by a cheater.[30][33]
Enhanced memory for faces of cheaters has been well demonstrated in humans.[30] Specifically, when exposed to trustworthy and untrustworthy faces, the memories of untrustworthy faces persist longer through an extinction procedure than do trustworthy faces, and when presented with pictures of faces with accompanying descriptions of cheating, trustworthy, or irrelevant information, people have enhanced source memory for the faces of cheaters.[22][34][35] However, the existence of a cognitive module specific to cheater detection is the subject of scientific debate.[30] While enhanced memory for cheaters is most likely due to a specific cognitive module,[22] there is currently insufficient evidence to support the existence of this specific module due to the difficulty of demonstrating such an evolutionary cognitive trait.[20][35]
Cheater recognition is closely related to the recognition of specific emotions.[30] Bell et al. argue for the existence of an emotional incongruity effect.[36] Memory is better for the face of a smiling cheater than of a smiling cooperator, because the incongruity captures the attention of the participant. If the emotion violates the expectancy then it is much better remembered. Furthermore, negative information is much better remembered whether it violates expectancies or not.[37] Negative emotions have a strong effect on source memory due to the presence of threatening stimuli.[30][38] Memory is determined by the emotional reaction of people at the encoding stage for both trustworthy and cheaters' facial expressions.[39] Furthermore, encoding levels process the recognition for traits and features of face recognition over expression of the face.[40] Memory is more readily remembered for emotionally involving information than for emotionally neutral information.[30][41][42]
Friends and foes are represented differently in the brain. The fusiform cortex, posterior cingulate gyrus, amygdala, and areas involved in motivational control were differentially activated as a function of previous social encounters. In general, these areas were more active when faces were perceived as foes rather than friends.[43]
Destination memory
Destination memory is the ability to remember information one has conveyed to others. Telling a story multiple times, unaware that listeners have heard the story before, is an example of destination memory failure. Destination memory is important for conversations because it allows people to recall what was already talked about.[44] Deficiency in destination memory is more common among older adults, but has been reported by university students as well.[44] Destination memory is notoriously poor due to ineffective integration of components of episodic memory.[44] However, there appear to be few, if any, negative social implications for poor destination memory.
Implicit memory
Implicit memory is the non-declarative, or unspoken, aspects of memory.[45] Priming, motor memory and classical conditioning are all examples of implicit memory. An example of implicit memory's effect on our social interactions has been illustrated by the pin-in-hand phenomenon.[46] This phenomenon was first observed by Claparède[46] while dealing with an amnesiac patient. Normally, he would shake the patient's hand and introduce himself, as the patient thought it was their first time meeting. However, on one occasion he hid a pin and pricked the patient with it when she reached to shake his hand. Later, when he offered his hand, she drew away and refused to shake, in spite of the fact she could not explicitly recall the pinprick itself.[46] The patient was unable to provide a reason for her hesitation.
Memory for social norms
Social norms are unspoken rules that govern acceptable behaviours in a given social situation. Violation of these norms can be considered extremely offensive, and may lead to social rejection. Thus, it is adaptive to be able to adhere to social norms, which requires memory of them. As a person matures, they tend to passively gain an intricate understanding of the norms associated with their culture.[47]
Schemata
One way memory can affect social interactions is through schemata. A schema is a mental structure implemented in order to sort knowledge into central themes or subjects.[48] Although schemata develop from experience, not all experiences are organized with schemata.[49] Schemata can influence the way in which new people and situations are understood, which has a direct effect on social interaction.[48]
Many research studies have been conducted concerning the effect of schemata on individuals' perceptions of others.[50] It has been found that preschoolers are aware of gender schemata and are capable of pointing out behaviours which are inconsistent with them.[51] In a Canadian study, participants were asked to rate various diverse groups based on how well a given characteristic applied to them.[50] The results confirmed that participants were quicker to assign a rating if the given characteristics fit the schema for that group.[50]
Schemata have been proposed as a way to explain the persistence of stereotypes. According to this theory, negative stereotypes persist because information that is incongruent with a schema is not organized and thus less likely to be remembered.[48] Therefore, schemata influence what information is encoded and retrieved from memory, thereby influencing social interactions.
Impairments
Any condition that causes a memory deficit will affect social functioning. This will differ based on the nature and severity of the deficit. For example, patients with Alzheimer's disease have difficulty during social interactions because of agnosia, and aphasia.[11]
See also
References
- ↑ Pascalis, O.; De Schonen, S.; Morton, J.; Deruelle, C.; Fabre-Grenet, M. (1995). "Mother's Face Recognition by Neonates: A Replication and an Extension". Infant Behavior and Development. 18: 79–85. doi:10.1016/0163-6383(95)90009-8.
- ↑ Morton, J.; Johnson, M.H. (1991). "CONSPEC and CONLERN: A Two-Process Theory of Infant Face Recognition". Psychological Review. 98 (2): 164–181. doi:10.1037/0033-295x.98.2.164. PMID 2047512.
- 1 2 3 Sergent, J.; Ohta, S.; MacDonald, B. (1992). "Functional neuroanatomy of face and object processing. A positron emission tomography study". Brain. 115 (1): 15–36. doi:10.1093/brain/115.1.15. PMID 1559150.
- ↑ Kanwisher, N.; McDermott, J.; Chun, M.M. (1997). "The Fusiform Face Area: A Module in Human Extrastriate Cortex Specialized for Face Perception". The Journal of Neuroscience. 17 (11): 4302–4311.
- ↑ Barton, J.S.; Press, J.P.; Keenan, D.P.; et al. (2002). "Lesions of the fusiform face area impair perception of facial configuration in prosopagnosia". Neurology. 58: 71–78. doi:10.1212/wnl.58.1.71.
- ↑ Amunts, K.; Kedo, O.; Kindler, M.; Pieperhoff, P.; Mohlberg, H.; Shah, N.; Habel, U.; Schneider, F.; Zilles, K. (2005). "Cytoarchitectonic mapping of the human amygdala, hippocampal region and entorhinal cortex: intersubject variability and probability maps". Anat Embryol. 210: 343–352. doi:10.1007/s00429-005-0025-5. PMID 16208455.
- ↑ Adolphs, R.; Baron-Cohen, S.; Tranel, D. (2002). "Impaired Recognition of Social Emotions following Amygdala Damage". Journal of Cognitive Neuroscience. 14 (8): 1264–1274. doi:10.1162/089892902760807258.
- 1 2 Adolphs, R.; Tranel, D.; Damasio, A.R. (1998). "The human amygdala in social judgment". Nature. 393: 470–474. doi:10.1038/30982. PMID 9624002.
- ↑ Squire, L.R.; Zola-Morgan, S. (1991). "The medial temporal lobe memory system". Science. 253 (5026): 1380–1386. doi:10.1126/science.1896849.
- ↑ Milner, B. (1966). Amnesia following operation on the temporal lobes. In C.W.M. Whitty & O.L. Zangwill(Eds), Amnesia, (109-133). London: Butterworths.
- 1 2 American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders. 4th, text revision (DSM-IV-TR) ed. 2000.
- ↑ Leader, G.; Loughnane, A.; McMoreland, C.; Reed, P. (2009). "The Effect of Stimulus Salience on Over-selectivity". Journal of Autism and Developmental Disorders. 39: 330–338. doi:10.1007/s10803-008-0626-y.
- ↑ McPartland, J.C.; Webb, S.B.; Keehn, B.; Dawson, G. (2011). "Patterns of Visual Attention to Faces and Objects in Autism Spectrum Disorder". Journal of Autism and Developmental Disorders. 41: 148–157. doi:10.1007/s10803-010-1033-8. PMC 3074360.
- 1 2 O'Connor, K.; Kirk, I. (2008). "Brief Report: Atypical Social Cognition and Social Behaviours in Autism Spectrum Disorder: A Different Way of Processing Rather than an Impairment". Journal of Autism and Developmental Disorders. 38: 1989–1997. doi:10.1007/s10803-008-0559-5.
- ↑ Damasio, A.R.; Damasio, H.; Van Hoesen, G.W. (1982). "Prosopagnosia: anatomical basis and behavioural mechanics". Neurology. 32 (4): 331–341. doi:10.1212/wnl.32.4.331.
- 1 2 Goulder, A.W. (1960). "The Norm of Reciprocity: A Preliminary Statement". American Sociological Association. 25 (2): 161–178.
- 1 2 Milinski, M.; Semmann, D.; Krambeck, H.J. (2002). "Reputation helps solve the 'tragedy of the commons'". Nature. 415: 424–426. doi:10.1038/415424a. PMID 11807552.
- 1 2 Ashlock, D., Rogers, N. (2009). The Impact of Long-Term Memory in the Iterated Prisoner's Dilemma. "Archived copy" (PDF). Archived from the original (PDF) on 2011-07-06. Retrieved 2011-03-11.
- ↑ Milinski, M.; Wedekind, C. (1998). "Working memory constrains human cooperation in the Prisoner's Dilemma". Proceedings of the National Academy of Sciences of the United States of America. 95 (23): 13755–13758. doi:10.1073/pnas.95.23.13755. PMC 24892.
- 1 2 Nairne, James S.; Josefa N. S. Pandeirada (30 January 2010). "Memory Functions". Corsini Encyclopedia of Psychology. doi:10.1002/9780470479216.corpsy0536. Retrieved 4 December 2013.
- ↑ Nairne, James S.; Joshua E. VanArsdall; Josefa N. S. Pandeirada (2012). "Adaptive memory: Enhanced location memory after survival processing". Journal of Experimental Psychology: Learning, Memory, and Cognition. 38 (2): 495–501. doi:10.1037/a0025728.
- 1 2 3 "Evolutionary Psychology: Applications and Criticisms". Encyclopedia of Cognitive Science. 2 Feb 2010. Retrieved 4 December 2013.
- ↑ McBride, Dawn M.; Thomas, Brandon J.; Zimmerman, Corinne (2 March 2013). "A test of the survival processing advantage in implicit and explicit memory tests". Memory & Cognition. 41 (6): 862–871. doi:10.3758/s13421-013-0304-y.
- 1 2 Espejo, E. F. (2003). "Prefrontocortical Dopamine Loss in Rats Delays Long-Term Extinction of Contextual Conditioned Fear, and Reduces Social Interaction Without Affecting Short-Term Social Interaction Memory". Neuropsychopharmacology. 28 (3): 490–498. doi:10.1038/sj.npp.1300066. PMID 12629528.
- 1 2 Belmore, S (1987). "Determinants of attention during impression formation". Journal of Experimental Psychology: Learning, Memory, and Cognition. 13 (3): 480–489. doi:10.1037/0278-7393.13.3.480.
- 1 2 3 Srull, T. K. (1981). "Person memory: Some tests of associative storage and retrieval models". Journal of Experimental Psychology: Human Learning and Memory. 7: 440–462. doi:10.1037/0278-7393.7.6.440.
- ↑ Hastie, R (1984). "Causes and effects of causal attribution". Journal of Personality and Social Psychology. 46: 44–56. doi:10.1037/0022-3514.46.1.44.
- 1 2 3 4 5 Ybarra, O.; Schaberg, L.; Keiper, S. (1999). "Favourable and Unfavourable Target Expectations and Social Information Processing". Journal of Personality and Social Psychology. 77 (4): 698–709. doi:10.1037/0022-3514.77.4.698.
- ↑ Skowronski, J.; Carlston, D. (1987). "Social Judgement and Social Memory: The Role of Cue Diagnosticity in Negativity, Positivity, and the Extremity Bias". Journal of Personality and Social Psychology. 52 (4): 689–699. doi:10.1037/0022-3514.52.4.689.
- 1 2 3 4 5 6 7 Bell, R.; Buchner, A. (3 December 2012). "How Adaptive Is Memory for Cheaters?". Current Directions in Psychological Science. 21 (6): 403–408. doi:10.1177/0963721412458525.
- ↑ Suzuki, Atsunobu; Honma, Suga (November 2013). "Indelible Distrust: Memory Bias Toward Cheaters Revealed as High Persistence Against Extinction". Journal of Experimental Psychology: Learning, Memory, and Cognition. 6. 39.
- ↑ Volstorf, Jenny; Rieskamp, Stevens (2011). "The Good, the Bad, and the Rare: Memory". PLoS ONE. 6 (4): e18945. doi:10.1371/journal.pone.0018945. PMC 3084729. PMID 21559490.
- ↑ Bell, R; Kroneisen, Giang (November 2012). "On the Flexibility of Social Source Memory: A Test of the Emotional Incongruity Hypothesis". Journal of Experimental Psychology: Learning, Memory, and Cognition. 38 (6).
- ↑ Buchner, Axel; Bell, Raoul; Mehl, Bettina; Musch, Jochen (May 2009). "No enhanced recognition memory, but better source memory for faces of cheaters". Evolution and Human Behavior. 30 (3): 212–224. doi:10.1016/j.evolhumbehav.2009.01.004.
- 1 2 Suzuki, Atsunobu; Honma, Yoshiko; Suga, Sayaka (2013). "Indelible distrust: Memory bias toward cheaters revealed as high persistence against extinction". Journal of Experimental Psychology: Learning, Memory, and Cognition. 39 (6): 1901–1913. doi:10.1037/a0033335.
- ↑ Bell, R.; Buchner, A.; Giang, T.; Kroneise, M. (2012). "On the flexibility of social source memory: A test of the emotional incongruity Hypothesis". Journal of Experimental Psychology: Learning, Memory, and Cognition. 38 (6): 1512–1529. doi:10.1037/a0028219.
- ↑ Ochsner, K. N. (2000). "Are affective events richly recollected or simply familiar? The experience and process of recognizing feelings past". Journal of Experimental Psychology: General. 129: 242–261. doi:10.1037/0096-3445.129.2.242. PMID 10868336.
- ↑ Bliss-Moure, E.; Barrett, L.F.; Wright, C. (2008). "Individual Differences in Learning the Affective Value of Others Under Minimal Conditions". Emotion. 8 (4): 479–493. doi:10.1037/1528-3542.8.4.479. PMC 2761963.
- ↑ Baumeister, R. F.; Bratslavsky, E.; Finkenauer, C.; Vohs, K. D. (2001). "Bad is stronger than good". Review of General Psychology. 5: 323–370. doi:10.1037/1089-2680.5.4.323.
- ↑ McKelvie, Stuart J. (1995). Encoding Oporations and Recognition Memory for Faces. Canadian Journal of Experimental Psychology/Revue canadienne de psychologie expérimentale, 49(4), Dec, 1995. 437-459.
- ↑ D'Argembeau, A.; Van der Linden, M. (2004). "Influence of affective meaning on memory for contextual information". Emotion. 4: 173–188. doi:10.1037/1528-3542.4.2.173.
- ↑ D'Argembeau, A.; Van der Linden, M. (2004). "Influence of affective meaning on memory for contextual information". Emotion. 4: 173–188. doi:10.1037/1528-3542.4.2.173. Cheating, or cheater face recognition, is more memorable if applicable.
- ↑ Vrticka, P.; Andersson, F.; Sander, D.; Vuilleumier, P. (2009). "Memory for friends or foes: the social context of past encounters with faces modulates their subsequent neural traces in the brain". Social Neuroscience. 4 (5): 384–401. doi:10.1080/17470910902941793.
- 1 2 3 Gopie, N.; MacLeod, C. (2009). "Destination Memory: Stop Me if I've Told You This Before". Psychological Science. 20 (12): 1492–1499. doi:10.1111/j.1467-9280.2009.02472.x. PMID 19891750.
- ↑ Schacter, D.L. (1987). "Implicit Memory: History and Current Status". Journal of Experimental Psychology. 13 (3): 501–518. doi:10.1037/0278-7393.13.3.501.
- 1 2 3 Feinstein, J.S., Duff, M.C., Tranel, D. (2010). Sustained experience of emotion after loss of memory in patients with amnesia. "Proceedings of the National Academy of Sciences of the United States of America". 107(17), 7674-7679.
- ↑ Fehr, E.; Fischbacher, U. (2003). "The nature of human altruism". Nature. 425: 785–791. doi:10.1038/nature02043. PMID 14574401.
- 1 2 3 Aronson. E., Wilson T.D., Akert R.M., Fehr, B., (2007) Social psychology: third canadian edition. Pearson Education Canada., TO, ON. p61.
- ↑ Marshall, S.P. (1995). Schemata in problem solving. NY, NY: Cambridge press.
- 1 2 3 Macrea, C.N., Stangor, C., & Hewstone, M. (1996). Stereotypes and stereotyping. New York, NY : The Guilford Press.
- ↑ Carter, D.B., & Levy, G.D. (1988). Cognitive aspects of early sex-role development. Child Development, 59. Retrieved from https://www.jstor.org/pss/1130576