Splicing
Splicing is in de genetica een verandering van genetische informatie na transcriptie. Tijdens de RNA-processing worden de introns uit het pre-mRNA geknipt en de exons van het pre-mRNA aan elkaar geplakt. Introns komen hoofdzakelijk voor in eukaryotische cellen. De term splicing is afgeleid van het splitsen van touwen, waarbij gedeelten van een touw samengevoegd worden door strengen in elkaar te draaien.
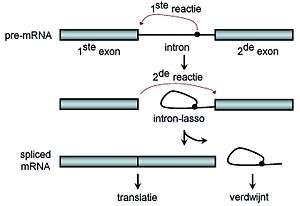
Als de introns uit het pre-mRNA geknipt zijn (dit heet cleavage), blijven enkel de exons over. Het verbinden van deze exons tot een nieuw geheel, het mRNA, heet splicing, na de capping (letterlijk kapje opzetten, is het toevoegen van 7-methylguanosine aan de kop) en polyadenylatie (poly-A-staart op het einde) is het mRNA klaar om de translatie te ondergaan.
Splicing, capping en polyadenylatie vinden tegelijkertijd tijdens de transcriptie plaats. Dit cotranscriptioneel RNA-proces wordt wel de "RNA-fabriek" genoemd.
Typen van splicing
Er komen in de natuur verschillende typen van splicing voor. De wijze van splicing hangt af van het intron en de daarbij behorende katalysator. Bij alle typen van splicing worden de uitgeknipte introns afgebroken.
Spliceosomaal
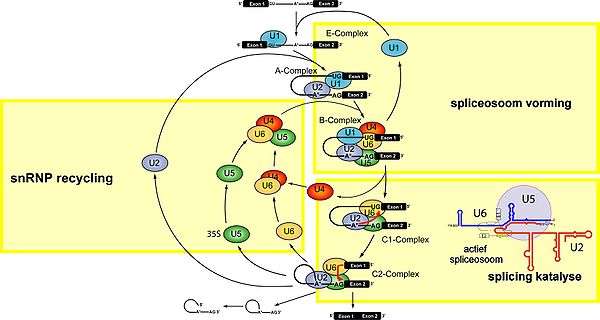
Spliceosomale introns worden vaak aangetroffen in eukaryotische, proteïnecoderende genen. Voor de splicing zijn een 3'-spliceplaats, 5'-spliceplaats en een zijtakplaats (branche site) op het intron noodzakelijk. Splicing wordt gekatalyseerd door het spliceosoom, dat bestaat uit een lang RNA-proteïnecomplex en is samengesteld uit vijf kleine celkern- (nucleaire) ribonucleoproteïnen (snRNP's, uitgesproken als "snurps"). Het RNA-gedeelte van de snRNP's reageert met het intron en is waarschijnlijk betrokken bij de katalyse. Er zijn twee typen spliceosomen bekend (de major en de minor) met verschillende snRNP's.
- Major
- Het majorspliceosoom splicet introns die GU (guanine, uracil) op de 5'-spliceplaats en AG (adenine, guanine) op de 3'-spliceplaats hebben zitten. Het bestaat uit de U1-, U2-, U4-, U5- en U6-snRNP's.
- Minor
- Het minorspliceosoom lijkt veel op het majorspliceosoom, maar het splicet zeldzaam voorkomende introns heeft verschillende volgorden (sequenties) van spliceplaatsen. Hierbij bestaan de 3'- en 5'-spliceplaatsen respectievelijk uit AU (adenine, uracil) en AC (adenine, cytosine). Het minorspliceosoom heeft evenals het major het U5-snRNP, maar andere snRNP's voor U1, U2, U4 en U6, die respectievelijk U11, U12, U4atac, en U6atac genoemd worden.
- Transsplicing
- Transsplicing is een splicingvorm die twee exons die niet in hetzelfde RNA-transcript zitten, met elkaar verbindt.
Zelfsplicing
Zelfsplicing vindt plaats bij sommige zeldzaam voorkomende introns, die een ribozym vormen. Hierbij wordt de splicing door het RNA alleen uitgevoerd, dus zonder spliceosomen. Er zijn drie groepen van zelfsplicing introns, Groep I, II en III. Groep II- en III-introns voeren de splicing vergelijkbaar met het spliceosoom uit, maar gebruiken geen proteïnekatalysator. Evolutionair gezien vertonen ze veel overeenkomst met het spliceosoom.
In 1981 kon een onderzoeksgroep onder leiding van Thomas Cech aantonen dat bij de voorloper van het 26S rRNA van Tetrahymena thermophila, voor de vorming van een ongeveer 400 nucleotide lang intron geen proteïne nodig was. Hiervoor kreeg Cech samen met Sidney Altman in 1989 de Nobelprijs voor de Scheikunde. Latere onderzoeken toonden aan dat zelfsplicing in veel meer organismen optreedt.
De Groep I-introns komen voor bij pre-mRNA's in eenvoudige eukaryoten, zoals Tetrahymena thermophila en bij pre-mRNA's in celorganen als mitochondriën en chloroplasten. Het uitknippen van de introns gebeurt in twee stappen. Als katalysator treedt guanosine op, die door de structuur van het RNA op de juiste manier gevouwen wordt, waardoor deze aan de 5'-spliceplaats een reactie doorvoert. De 3'-hydroxygroep van het 5'-eind van het exon maakt verbinding met de 3'-spliceplaats van het volgende exon, waardoor de beide exons aan elkaar gaan zitten en tegelijkertijd het intron vrijkomt. Het intron vormt uiteindelijk een ring.
De Groep II-introns komen voor bij pre-mRNA's in de mitochondriën van gisten en andere schimmels. Een guanosinecofactor is hierbij niet nodig. Door de structuur van het RNA wordt het adenosine, dat 7 of 8 nucleotiden vanaf de 3'-spliceplaats ligt, zodanig gestructureerd dat de 5'-spliceplaats van het nucleofiel met zijn 2'-hydroxygroep aan het adenosine kan vastmaken. Hierdoor ontstaat een ongewone 2'-5'-fosfodiesterbinding en daarmee een lassovormig intron (lariat). In de tweede stap, die vergelijkbaar is met die van Groep I, worden de exons met elkaar verbonden en komen de introns vrij.
tRNA splicing
tRNA (ook tRNA-achtige) splicing is een andere zeldzaam voorkomende vorm van splicing die gewoonlijk optreedt in tRNA. Het komt zowel voor bij Archaea als bij eukaryoten. Hierbij zijn de biochemische reacties anders dan bij spliceomsomaal en zelfsplicing. Ribonucleasen knippen het RNA, ligasen plakken de exons aan elkaar.
Bij deze enzymatische splicing worden de introns niet door hun sequentie herkend, maar door een structuur die om het gehele molecuul zit, zoals het bulge-helix-bulgemotief (BHB-motief) bij Archaea, en worden ze in drie stappen verwijderd. Het pre-tRNA wordt tweemaal door een endonuclease geknipt, waardoor het intron en twee zogenoemde tRNA-halfmoleculen vrijkomen. Het daarbij gevormde cyclische 2'-3'-fosfaat van het 5'-halfmolecuul worden gehydrolyseerd tot een 2'-fosfaat en een 3'-OH-groep, waarbij de 5'-OH-groep van het 3'-halfmolecuul met verbruik van GTP gefosforyliseerd wordt. Daardoor wordt ligatie met een RNA-ligase en ATP-hydrolyse mogelijk. Ten slotte wordt in de laatste stap het 2'-fosfaat verwijderd met verbruik van NAD en de vorming van nicotinamide. Er zijn ook enkele mRNA's bekend, waarbij hetzelfde proces als bij tRNA's optreedt.
Biochemisch mechanisme
Spliceosomale splicing en zelfsplicing gebeurt in twee stappen, waarbij in beide stappen transesterificatiereacties tussen RNA-nucleotiden optreden. Bij tRNA-splicing treedt geen transesterificatie op. De transesterificatiereacties vinden in een bepaalde volgorde plaats. Allereerst vormt een nucleotide op de zijtakplaats (branche site) met de voorste nucleotide van het intron een "intronlasso" (lariat). Daarna reageert de achterste nucleotide van het eerste exon met de voorste nucleotide van het tweede exon en verbindt zo de twee exons met elkaar, waarbij de intronlasso loslaat en verdwijnt.
Alternatieve splicing
In veel gevallen kunnen door variaties in het splicingproces veel verschillende proteïnen met hetzelfde mRNA gevormd worden: de zogenaamde proteïne isovormen. Dit proces wordt alternatieve splicing genoemd.
Kunstmatige splicing
Het is mogelijk door kunstgrepen het splicingproces te beïnvloeden.
Splicingfouten
Mutaties in de introns of exons kunnen de splicing blokkeren, waardoor er geen eiwitsynthese kan plaatsvinden.
Voorkomende fouten:
- Mutatie van een spliceplaats, waardoor deze niet meer kan functioneren en resulteert in verlies van een exon of het niet uitknippen van een intron.
- Mutatie van een spliceplaats, waardoor verminderde specificiteit optreedt. Dit geeft verschil in spliceplaats, waardoor insertie of deletie van aminozuren of helemaal geen vorming van aminozuren optreedt.
- Transpositie (verplaatsing) van een spliceplaats, waardoor lange of korte exons gevormd worden.
Aandoeningen
Een klassiek voorbeeld van een aandoening die het gevolg is van een fout bij splicing, is β-thalassemie. Het is een erfelijke aandoening, die ontstaan is door een puntmutatie van het gen op de 5' spliceplaats van intron 1. Hierdoor vormt het spliceosoom korte of lange mRNA’s, die vervolgens onbruikbare proteïnen aanmaken.
Een ander voorbeeld is een mutatie in intron 2 van hetzelfde gen, waardoor een kort stukje van het intron 2 in het mRNA blijft zitten.
In beide gevallen is de hemoglobinesynthese in de erytrocyten sterk geremd en is er onvoldoende zuurstoftransport mogelijk.
Het syndroom van Ehlers-Danlos (EDS) Type II is het gevolg van een mutatie in de zijtakplaats (branche site) van het gen COL5A1.
Spinale musculaire atrofie (SMA) is het gevolg van een mutatie in de splicing-enhancers/silencers van het gen SMN1 (enhancing is de mogelijkheid van het aflezen en bij silencing is aflezen niet mogelijk).
"RNA-fabriek" (RNA-factory)
De verschillende processen, zoals transcriptie, mRNA-vorming, mRNA-transport naar het cytoplasma, RNA-lokalisering, translatie en RNA-afbraak, beïnvloeden en reguleren elkaar. De vorming van pre-mRNA blijft ook tijdens het splicingproces doorgaan. Daarom spreekt men ook wel van de "RNA-fabriek" (RNA-factory).
Splicing, dat in de celkern plaatsvindt, kan zelfs de processen buiten de celkern (in het cytoplasma) beïnvloeden. Een proteïnecomplex wordt door het spliceosoom aan het rijpe mRNA vastgehecht (het Exon-Junction-Complex, EJC), waardoor er een effectief transport van het mRNA uit de celkern kan plaatsvinden. Hierbij wordt tegelijkertijd extra informatie overgedragen, die voor een latere kwaliteitscontrole van het mRNA tijdens de translatie zorg draagt (Nonsense-mediated mRNA decay, NMD). Een volledig pre-mRNA komt in een cel niet voor, omdat de introns al tijdens de transcriptie worden verwijderd.
Overzicht van alle processen
- DNA — transcriptie → pre-mRNA — RNA-processing, splicing, RNA-editing of alternatieve splicing → mRNA — translatie → eiwit