Tetrachromacy
Tetrachromacy is the condition of possessing four independent channels for conveying color information, or possessing four types of cone cell in the eye. Organisms with tetrachromacy are called tetrachromats.
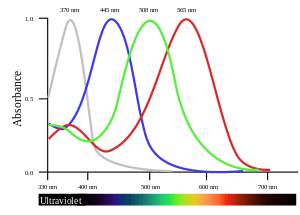
In tetrachromatic organisms, the sensory color space is four-dimensional, meaning that to match the sensory effect of arbitrarily chosen spectra of light within their visible spectrum requires mixtures of at least four primary colors.
Tetrachromacy is demonstrated among several species of bird, fish, amphibian, reptile, insect and some mammals.[2][3] It was the normal condition of most mammals in the past; a genetic change made the majority of species of this class eventually lose two of their four cones.[4][5]
Physiology
The normal explanation of tetrachromacy is that the organism's retina contains four types of higher-intensity light receptors (called cone cells in vertebrates as opposed to rod cells, which are lower-intensity light receptors) with different absorption spectra. This means that the organism may see wave-lengths beyond those of a typical human's vision, and may be able to distinguish between colors that, to a normal human, appear to be identical. Species with tetrachromatic color vision may have an unknown physiological advantage over rival species.[6]
Examples
Fish
The goldfish (Carassius auratus auratus)[7] and zebrafish (Danio rerio)[8] are examples of tetrachromats, containing cone cells sensitive for red, green, blue and ultraviolet light.
Birds
Some species of birds, such as the zebra finch and the Columbidae, use the ultraviolet wave-length 300–400 nm specific to tetrachromatic color vision as a tool during mate selection and foraging.[9] When selecting for mates, ultraviolet plumage and skin coloration show a high level of selection.[10] A typical bird eye will respond to wave-lengths from about 300 to 700 nm. In terms of frequency, this corresponds to a band in the vicinity of 430–1000 THz. Most birds have retinas with four spectral types of cone cell that are believed to mediate tetrachromatic color vision. Bird color vision is further improved by the filtering of pigmented oil droplets that are located in the photoreceptors. The oil droplets filter incident light before it reaches the visual pigment in outer segments of the photoreceptors.
The four cone types, and the specialization of pigmented oil droplets, give birds better color vision than that of humans .[11][12] However, more recent research has suggested that tetrachromacy in birds only provide birds with a larger visual spectrum than that in humans (humans cannot see ultraviolet light, 300-400 nm), while the spectral resolution (the "sensitivity" to nuances) is similar.[13]
Insects
Foraging insects can see wave-lengths that flowers reflect (ranging from 300 nm to 700 nm[14][15]). Pollination being a mutualistic relationship, foraging insects and some plants have coevolved, both increasing wave-length range: in perception (pollinators), in reflection and variation (flower colors).[6] Directional selection has led plants to display increasingly diverse amounts of color variations extending into the ultraviolet color scale, thus attracting higher levels of pollinators.[6]
Mammals
Reindeer
In areas where reindeer live, the sun remains very low in the sky for long periods. Some parts of the environment absorb ultraviolet light and therefore to UV-sensitive reindeer, appear to be black, strongly contrasting with the snow. These include urine (indicating predators or competitors), lichens (a food source) and fur (as possessed by wolves, predators of reindeer).[16] Although reindeer do not possess a specific UV opsin, retinal responses to 330 nm have been recorded, mediated by other opsins.[17] It has been proposed that UV flashes on power lines are responsible for reindeer avoiding power lines because "...in darkness these animals [reindeer] see power lines not as dim, passive structures but, rather, as lines of flickering light stretching across the terrain."[18]
Humans
Apes (including humans) and Old World monkeys normally have three types of cone cell and are therefore trichromats. However, at low light intensities, the rod cells may contribute to color vision, giving a small region of tetrachromacy in the color space;[19] human rod cells' sensitivity is greatest at a bluish-green wave-length.
In humans, two cone cell pigment genes are present on the X chromosome: the classical type 2 opsin genes OPN1MW and OPN1MW2. People with two X chromosomes could possess multiple cone cell pigments, perhaps born as full tetrachromats who have four simultaneously functioning kinds of cone cell, each type with a specific pattern of responsiveness to different wave-lengths of light in the range of the visible spectrum.[20] One study suggested that 15% of the world's women might have the type of fourth cone whose sensitivity peak is between the standard red and green cones, giving, theoretically, a significant increase in color differentiation.[21] Another study suggests that as many as 50% of women and 8% of men may have four photopigments and corresponding increased chromatic discrimination compared to trichromats.[22] In 2010, after twenty years of study of women with four types of cones (non-functional tetrachromats), neuroscientist Dr. Gabriele Jordan identified a woman (subject cDa29) who could detect a greater variety of colors than trichromats could, corresponding with a functional tetrachromat (or true tetrachromat).[23][24][25][26]
Variation in cone pigment genes is wide-spread in most human populations, but the most prevalent and pronounced tetrachromacy would derive from female carriers of major red/green pigment anomalies, usually classed as forms of "color blindness" (protanomaly or deuteranomaly). The biological basis for this phenomenon is X-inactivation of heterozygotic alleles for retinal pigment genes, which is the same mechanism that gives the majority of female new-world monkeys trichromatic vision.[27]
In humans, preliminary visual processing occurs in the neurons of the retina. It is not known how these nerves would respond to a new color channel, that is, whether they could handle it separately or just combine it in with an existing channel. Visual information leaves the eye by way of the optic nerve; it is not known whether the optic nerve has the spare capacity to handle a new color channel. A variety of final image processing takes place in the brain; it is not known how the various areas of the brain would respond if presented with a new color channel.
Mice, which normally have only two cone pigments, can be engineered to express a third cone pigment, and appear to demonstrate increased chromatic discrimination,[28] arguing against some of these obstacles; however, the original publication's claims about plasticity in the optic nerve have also been disputed.[29]
Humans cannot see ultraviolet light directly because the lens of the eye blocks most light in the wavelength range of 300–400 nm; shorter wavelengths are blocked by the cornea.[30] The photoreceptor cells of the retina are sensitive to near ultraviolet light, and people lacking a lens (a condition known as aphakia) see near ultraviolet light (down to 300 nm) as whitish blue, or for some wavelengths, whitish violet, probably because all three types of cones are roughly equally sensitive to ultraviolet light; however, blue cone cells are slightly more sensitive.[31]
Tetrachromacy may also enhance vision in dim lighting, or when looking at a screen.[25]
See also
- Dichromacy
- Evolution of color vision
- Monochromacy
- Pentachromacy
- RG color space
- RGBY
- Somatosensory amplification
- Supertaster
- Trichromacy
| Look up tetrachromacy in Wiktionary, the free dictionary. |
References
- Figure data, uncorrected absorbance curve fits, from Hart, NS; Partridge, JC; Bennett, ATD; Cuthill, IC (2000). "Visual pigments, cone oil droplets and ocular media in four species of estrildid finch". Journal of Comparative Physiology A. 186 (7–8): 681–694. doi:10.1007/s003590000121. PMID 11016784.
- Goldsmith, Timothy H. (2006). "What Birds See". Scientific American (July 2006): 69–75.
- Wilkie, Susan E.; Vissers, Peter M. A. M.; Das, Debipriya; Degrip, Willem J.; Bowmaker, James K.; Hunt, David M. (1998). "The molecular basis for UV vision in birds: spectral characteristics, cDNA sequence and retinal localization of the UV-sensitive visual pigment of the budgerigar (Melopsittacus undulatus)". Biochemical Journal. 330 (Pt 1): 541–47. doi:10.1042/bj3300541. PMC 1219171. PMID 9461554.
- Jacobs, G. H. (2009). "Evolution of colour vision in mammals". Phil. Trans. R. Soc. B. 364 (1531): 2957–2967. doi:10.1098/rstb.2009.0039. PMC 2781854. PMID 19720656.
- Arrese, C. A.; Runham, P. B; et al. (2005). "Cone topography and spectral sensitivity in two potentially trichromatic marsupials, the quokka (Setonix brachyurus) and quenda (Isoodon obesulus)". Proc. Biol. Sci. 272 (1565): 791–796. doi:10.1098/rspb.2004.3009. PMC 1599861. PMID 15888411.
- Backhaus, W., Kliegl, R., Werner, J.S. (1998). "Color vision: perspective from different disciplines": 163–182. Cite journal requires
|journal=(help)CS1 maint: multiple names: authors list (link) - Neumeyer, Christa (1988). Das Farbensehen des Goldfisches: Eine verhaltensphysiologische Analyse. G. Thieme. ISBN 978-3137187011.
- Robinson, J.; Schmitt, E.A.; Harosi, F.I.; Reece, R.J.; Dowling, J.E. (1993). "Zebrafish ultraviolet visual pigment: absorption spectrum, sequence, and localization". Proc. Natl. Acad. Sci. U.S.A. 90 (13): 6009–6012. Bibcode:1993PNAS...90.6009R. doi:10.1073/pnas.90.13.6009. PMC 46856. PMID 8327475.
- Bennett, Andrew T. D.; Cuthill, Innes C.; Partridge, Julian C.; Maier, Erhard J. (1996). "Ultraviolet vision and mate choice in zebra finches". Nature. 380 (6573): 433–435. Bibcode:1996Natur.380..433B. doi:10.1038/380433a0.
- Bennett, Andrew T. D.; Théry, Marc (2007). "Avian Color Vision and Coloration: Multidisciplinary Evolutionary Biology". The American Naturalist. 169 (S1): S1–S6. doi:10.1086/510163. ISSN 0003-0147. JSTOR 510163.
- Cuthill, Innes C.; Partridge, Julian C.; Bennett, Andrew T. D.; Church, Stuart C.; Hart, Nathan S.; Hunt, Sarah (2000). J. B. Slater, Peter; Rosenblatt, Jay S.; Snowdon, Charles T.; Roper, Timothy J. (eds.). Ultraviolet Vision in Birds. Advances in the Study of Behavior. 29. Academic Press. p. 159. doi:10.1016/S0065-3454(08)60105-9. ISBN 978-0-12-004529-7.
- Vorobyev, M. (November 1998). "Tetrachromacy, oil droplets and bird plumage colours". Journal of Comparative Physiology A. 183 (5): 621–33. doi:10.1007/s003590050286. PMID 9839454.
- Olsson, Peter; Lind, Olle; Kelber, Almut (2015-01-15). "Bird colour vision: behavioural thresholds reveal receptor noise". Journal of Experimental Biology. 218 (2): 184–193. doi:10.1242/jeb.111187. ISSN 0022-0949. PMID 25609782.
- Markha, K. R.; Bloor, S. J.; Nicholson, R.; Rivera, R.; Shemluck, M.; Kevan, P. G.; Michener, C. (2004). "Black flower coloration in wild lisianthius nigrescens". Z Naturforsch C. 59c (9–10): 625–630. doi:10.1515/znc-2004-9-1003. PMID 15540592.
- Backhaus, W.; Kliegl, R.; Werner, J. S., eds. (1998). Colour Vision: Perspectives from Different Disciplines. pp. 45–78. ISBN 9783110161007.
- "Reindeer use UV light to survive in the wild". UCL News. May 26, 2011. Retrieved May 25, 2016.
- Hogg, C., Neveu, M., Stokkan, K.A., Folkow, L., Cottrill, P., Douglas, R., ... and Jeffery, G. (2011). "Arctic reindeer extend their visual range into the ultraviolet" (PDF). Journal of Experimental Biology. 214 (12): 2014–2019. doi:10.1242/jeb.053553. PMID 21613517.CS1 maint: multiple names: authors list (link)
- Tyler, N., Stokkan, K.A., Hogg, C., Nellemann, C., Vistnes, A.I. and Jeffery, G. (2014). "Ultraviolet vision and avoidance of power lines in birds and mammals". Conservation Biology. 28 (3): 630–631. doi:10.1111/cobi.12262. PMC 4232876. PMID 24621320.CS1 maint: multiple names: authors list (link)
- Hansjochem Autrum & Richard Jung (1973). Integrative Functions and Comparative Data. 7. Springer-Verlag. p. 226. ISBN 978-0-387-05769-9.
- Jameson, K. A., Highnote, S. M., & Wasserman, L. M. (2001). "Richer color experience in observers with multiple photopigment opsin genes" (PDF). Psychonomic Bulletin and Review. 8 (2): 244–261. doi:10.3758/BF03196159. PMID 11495112. Archived from the original (PDF) on 14 February 2012.CS1 maint: multiple names: authors list (link)
- Jordon, G. (July 1993). "A study of women heterozygous for colour deficiencies". Elsevier.
- Backhaus, Werner G. K.; Backhaus, Werner; Kliegl, Reinhold; Werner, John Simon (1998). Color Vision: Perspectives from Different Disciplines. Walter de Gruyter. ISBN 9783110161007.
- Didymus, JohnThomas (Jun 19, 2012), "Scientists find woman who sees 99 million more colors than others", Digital Journal
- Jordan, Gabriele; Deeb, Samir S.; Bosten, Jenny M.; Mollon, J. D. (July 2010). "The dimensionality of color vision in carriers of anomalous trichromacy". Journal of Vision. 10 (12): 12. doi:10.1167/10.8.12. PMID 20884587.
- Robson, David (September 5, 2014). "The Women with Superhuman Vision". BBC News. Archived from the original on September 13, 2014. Retrieved December 30, 2017.
- San Diego woman Concetta Antico diagnosed with 'super vision' Published on 22 Nov 2013
- Richard C. Francis (2011). "Chapter 8. X-Women". Epigenetics: The Ultimate Mystery of Inheritance. New York and London: W. W. Norton. pp. 93–104. ISBN 978-0-393-07005-7.
- Jacobs, Gerald H.; Williams, Gary A.; Cahill, Hugh; Nathans, Jeremy (23 March 2007). "Emergence of Novel Color Vision in Mice Engineered to Express a Human Cone Photopigment". Science. 315 (5819): 1723–1725. Bibcode:2007Sci...315.1723J. doi:10.1126/science.1138838. PMID 17379811.
- Makous, W. (12 October 2007). "Comment on 'Emergence of Novel Color Vision in Mice Engineered to Express a Human Cone Photopigment'". Science. 318 (5848): 196. Bibcode:2007Sci...318..196M. doi:10.1126/science.1146084. PMID 17932271.
- M A Mainster (2006). "Violet and blue light blocking intraocular lenses: photoprotection versus photoreception". British Journal of Ophthalmology. 90 (6): 784–792. doi:10.1136/bjo.2005.086553. PMC 1860240. PMID 16714268.
- Hambling, David (29 May 2002). "Let the light shine in". The Guardian.
External links
- Goldsmith, Timothy H. "What Birds See" Scientific American July 2006. An article about the tetrachromatic vision of birds
- Thompson, Evan (2000). "Comparative color vision: Quality space and visual ecology." In Steven Davis (Ed.), Color Perception: Philosophical, Psychological, Artistic and Computational Perspectives, pp. 163–186. Oxford: Oxford University Press.
- Looking for Madam Tetrachromat By Glenn Zorpette. Red Herring magazine, 1 November 2000
- "Exploring the fourth dimension". University of Bristol School of Biological Sciences. March 20, 2009.
- Colors - The Perfect Yellow By Radiolab, 21 May 2012 (Explores tetrachromacy in humans)
- The dimensionality of color vision in carriers of anomalous trichromacy--Gabriele Jordan et al--Journal of Vision August 12, 2010:
- On Tetrachromacy Ágnes Holba & B. Lukács
- San Diego woman Concetta Antico diagnosed with 'super vision' San Diego ABC television report, November 22, 2013.
Color topics | ||||||||
|---|---|---|---|---|---|---|---|---|
| ||||||||
| Color science |
|  | ||||||
| Color philosophy |
| |||||||
| Color terms |
| |||||||
| Color organizations |
| |||||||
| Lists |
| |||||||
| Related |
| |||||||
| ||||||||