Treadmilling
Treadmilling is a phenomenon observed in many cellular cytoskeletal filaments, especially in actin filaments and microtubules. It occurs when one end of a filament grows in length while the other end shrinks resulting in a section of filament seemingly "moving" across a stratum or the cytosol. This is due to the constant removal of the protein subunits from these filaments at one end of the filament while protein subunits are constantly added at the other end.[1]
Detailed process
Dynamics of the filament
The cytoskeleton is a highly dynamic part of a cell and cytoskeletal filaments constantly grow and shrink through addition and removal of subunits. Directed crawling motion of cells such as macrophages relies on directed growth of actin filaments at the cell front (leading edge).
Microfilaments
The two ends of an actin filament differ in their dynamics of subunit addition and removal. They are thus referred to as the plus end (with faster dynamics, also called barbed end) and the minus end (with slower dynamics, also called pointed end).[2] This difference results from the fact that subunit addition at the minus end requires a conformational change of the subunits.[3] Note that each subunit is structurally polar and has to attach to the filament in a particular orientation.[4] As a consequence, the actin filaments are also structurally polar.
Elongating the actin filament occurs when free-actin (G-actin) bound to ATP associates with the filament. Under physiological conditions, it is easier for G-actin to associate at the positive end of the filament, and harder at the negative end.[5] However, it is possible to elongate the filament at either end. Association of G-actin into F-actin is regulated by the critical concentration outlined below. Actin polymerization can further be regulated by profilin and cofilin.[5] Cofilin functions by binding to ADP-actin on the negative end of the filament, destabilizing it, and inducing depolymerization. Profilin induces ATP binding to G-actin so that it can be incorporated onto the positive end of the filament.
Microtubules
Two main theories exist on microtubule movement within the cell: dynamic instability and treadmilling.[6] Dynamic instability occurs when the microtubule assembles and disassembles at one end only, while treadmilling occurs when one end polymerizes while the other end disassembles. However, the biological significance of treadmilling in vivo is not well characterized.[7] This is due to the fact that within a living cell, many microtubules are tightly anchored at one end of the filament. Some research has suggested that the differences in critical concentration between the positive and the negative end may be a way for the cell to prevent unwanted polymerization events.[7]
Critical concentration
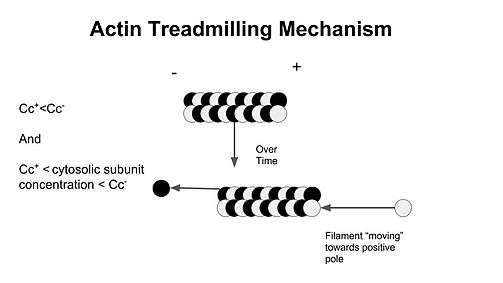
The critical concentration is the concentration of either G-actin (actin) or the alpha,beta- tubulin complex (microtubules) at which the end will remain in an equilibrium state with no net growth or shrinkage.[5] What determines whether the ends grow or shrink is entirely dependent on the cytosolic concentration of available monomer subunits in the surrounding area.[8] Critical concentration differs from the positive (CC+) and the negative end (CC−), and under normal physiological conditions, the critical concentration is lower at the positive end than the negative end. Examples in how the cytosolic concentration relates to the critical concentration and polymerization are as follows:
- A cytosolic concentration of subunits above both the CC+ and CC− ends results in subunit addition at both ends
- A cytosolic concentration of subunits below both the CC+ and CC− ends results in subunit removal at both ends
Note that the cytosolic concentration of the monomer subunit between the CC+ and CC− ends is what is defined as treadmilling in which there is growth at the plus end, and shrinking on the minus end.
The cell attempts to maintain a subunit concentration between the dissociation constants at the plus and minus ends of the polymer.
Steady-state treadmilling
While treadmilling may occur at different speeds at both ends, there is a concentration at which the speed of growth at the (+) end is equal to the rate of shrinkage at the (-) end. This is deemed steady-state treadmilling in which the net length of the treadmilling filament remains unchanged.
References
- Bruce Alberts, Dennis Bray, Julian Lewis: Molecular Biology of the Cell, 4th Edition, Taylor & Francis, 2002, pp. 909-920, ISBN 0-8153-4072-9
- Bruce Alberts (2008). Molecular biology of the cell. Garland Science. ISBN 978-0-8153-4105-5. Retrieved 4 February 2012.
- Alberts, B; Johnson, A; Lewis, J; et al. (2002). The Self-Assembly and Dynamic Structure of Cytoskeletal Filaments. Garland Science. Retrieved 19 October 2015.
- Gardet, A; Breton, M; Trugnan, G; Chwetzoff, S (2007). "Role for actin in the polarized release of rotavirus". Journal of Virology. 81 (9): 4892–4. doi:10.1128/JVI.02698-06. PMC 1900189. PMID 17301135.
- Remedios, C. G. Dos; Chhabra, D.; Kekic, M.; Dedova, I. V.; Tsubakihara, M.; Berry, D. A.; Nosworthy, N. J. (2003-04-01). "Actin Binding Proteins: Regulation of Cytoskeletal Microfilaments". Physiological Reviews. 83 (2): 433–473. doi:10.1152/physrev.00026.2002. ISSN 0031-9333. PMID 12663865.
- Rodionov, Vladimir I.; Borisy, Gary G. (1997-01-10). "Microtubule Treadmilling in Vivo". Science. 275 (5297): 215–218. doi:10.1126/science.275.5297.215. ISSN 0036-8075. PMID 8985015.
- Kirschner, MW (1980-07-01). "Implications of treadmilling for the stability and polarity of actin and tubulin polymers in vivo". The Journal of Cell Biology. 86 (1): 330–334. doi:10.1083/jcb.86.1.330. ISSN 0021-9525. PMC 2110666. PMID 6893454.
- Schaus, T. E.; Taylor, E. W.; Borisy, G. G. (2007). "Self-organization of actin filament orientation in the dendritic-nucleation/array-treadmilling model". Proceedings of the National Academy of Sciences. 104 (17): 7086–7091. Bibcode:2007PNAS..104.7086S. doi:10.1073/pnas.0701943104. PMC 1855413. PMID 17440042.