Pronucleus
A pronucleus (plural: pronuclei) is the nucleus of a sperm or an egg cell during the process of fertilization. The sperm cell becomes a pronucleus after the sperm enters the ovum, but before the genetic material of the sperm and egg fuse. Contrary to the sperm cell, the egg cell has a pronucleus once it becomes haploid, and not when the sperm cell arrives. Sperm and egg cells are haploid, meaning they carry half the number of chromosomes of somatic cells, so in humans, haploid cells have 23 chromosomes, while somatic cells have 46 chromosomes. The male and female pronuclei don't fuse, although their genetic material does. Instead, their membranes dissolve, leaving no barriers between the male and female chromosomes. Their chromosomes can then combine and become part of a single diploid nucleus in the resulting embryo, containing a full set of chromosomes.
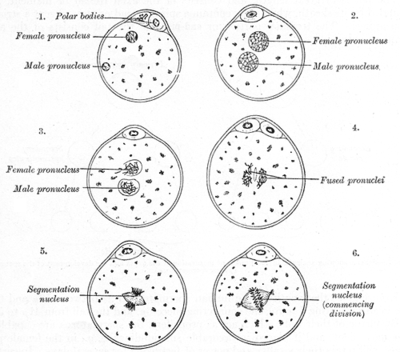
The appearance of two pronuclei is the first sign of successful fertilization as observed during in vitro fertilisation, and is usually observed 18 hours after insemination or ICSI. The zygote is then termed a two-pronuclear zygote (2PN). Two-pronuclear zygotes transitioning through 1PN or 3PN states tend to develop into poorer-quality embryos than ones who remain 2PN throughout development,[1] and may be significant in embryo selection in IVF.
History
The pronucleus was discovered the 1870s microscopically using staining techniques combined with microscopes with improved magnification levels. The pronucleus was originally found during the first studies on meiosis. Edouard Van Beneden published a paper in 1875 in which he first mentions the pronucleus by studying the eggs of rabbits and bats. He stated that the two pronuclei form together in the center of the cell to form the embryonic nucleus. Van Beneden also found that the sperm enters into the cell through the membrane in order to form the male pronucleus. In 1876, Oscar Hertwig did a study on sea urchin eggs because the eggs of sea urchins are transparent, so it allowed for much better magnification of the egg. Hertwig confirmed Van Beneden's finding of the pronucleus, and also found the formation of the female pronucleus involves the formation of polar bodies.[2]
Formation
The female pronucleus is the female egg cell once it has become a haploid cell, and the male pronucleus forms when the sperm enters into the female egg. While the sperm develops inside of the male testes, the sperm does not become a pronucleus until it decondenses quickly inside of the female egg.[3] When the sperm reaches the female egg, the sperm loses its outside membrane as well as its tail. The sperm does this because the membrane and the tail are no longer needed by the female ovum. The purpose of the cell membrane was to protect the DNA from the acidic vaginal fluid, and the purpose of the tail of the sperm was to help move the sperm cell to the egg cell. The formation of the female egg is asymmetrical, while the formation of the male sperm is symmetrical. Typically in a female mammal, meiosis starts with one diploid cell and becomes one haploid ovum and typically two polar bodies, however one may later divide to form a third polar body.[4] In a male, meiosis starts with one diploid cell and ends with four sperm.[5] In mammals, the female pronucleus starts in the center of the egg before fertilization. When the male pronucleus is formed, after the sperm cell reaches the egg, the two pronuclei migrate towards each other. However, in brown alga Pelvetia, the egg pronucleus starts in the center of the egg before fertilization and remain in the center after fertilization. This is because the egg cells of brown alga Pelvetia, the egg pronucleus is anchored down by microtubules so only the male pronucleus migrates towards the female pronucleus.[6]
Calcium Concentration
The calcium concentration within the egg cell cytoplasm has a very important role in the formation of an activated female egg. If there is no calcium influx, the female diploid cell will produce three pronuclei, rather than only one. This is due to the failure of release of the second polar body.[7]
Combination of Male and Female Pronuclei
The formation of the zygote starts with the fusion of both the inner and outer nuclei of the male and female pronuclei. It is unknown if one of the pronuclei start the combination of the two, or if the microtubules that help the dissolution of membranes commence the action.[8] The microtubules that make the two pronuclei combine come from the sperm’s centrosome. There is a study that strongly supports that microtubules are an important part of the fusion of the pronuclei. Vinblastine is a chemotherapy drug that affects both the plus and minus ends of microtubules.[9] When Vinblastine is added to the ovum, there is a high rate of pronuclear fusion failure. This high rate of pronuclear fusion failure highly suggests that microtubules play a major role in the fusion of the pronucleus.[10] In mammals, the pronuclei only last in the cell for about twelve hours, due to the fusion of the genetic material of the two pronuclei within the egg cell. Many studies of pronuclei have been in the egg cells of sea urchins, where the pronuclei are in the egg cell for less than an hour. The main difference between the process of fusion of genetic materials in mammals versus sea urchins is that in sea urchins, the pronuclei go directly into forming a zygote nucleus. In mammalian egg cells, the chromatin from the pronuclei form chromosomes that merge onto the same mitotic spindle. The diploid nucleus in mammals is first seen at the 2-cell stage, whereas in sea urchins it is first found at the zygote stage.[3]
References
- Reichman DE, Jackson KV, Racowsky C (May 2009). "Incidence and development of zygotes exhibiting abnormal pronuclear disposition after identification of two pronuclei at the fertilization check". Fertil. Steril. 94 (3): 965–970. doi:10.1016/j.fertnstert.2009.04.018. PMID 19476942.
- HAMOIR, GABRIEL. “The Discovery of Meiosis by E. Van Beneden, a Breakthrough in the Morphological Phase of Heredity.” Int. J. Dev. Biol., vol. 36, 1992, pp. 9–15.
- Gilbert SF. Developmental Biology. 6th edition. Sunderland (MA): Sinauer Associates; 2000. Fusion of the Genetic Material.
- Schmerler, Samuel; Wessel, Gary (January 2011). "Polar Bodies – more a lack of understanding than a lack of respect". Molecular Reproduction and Development. 78 (1): 3–8. doi:10.1002/mrd.21266. ISSN 1040-452X. PMC 3164815. PMID 21268179.
- Root Gorelick. “Meiosis Is Not Gender Neutral.” BioScience, vol. 62, no. 7, 2012, pp. 623–624. JSTOR, www.jstor.org/stable/10.1525/bio.2012.62.7.2.
- Swope, Richard E., and Darryl L. Kropf. “Pronuclear Positioning and Migration during Fertilization in Pelvetia.” Developmental Biology, vol. 157, no. 1, 1993, pp. 269–276., doi:10.1006/dbio.1993.1131.
- Miao, Yi-Liang, et al. “Calcium Influx-Mediated Signaling Is Required for Complete Mouse Egg Activation.” Proceedings of the National Academy of Sciences of the United States of America, vol. 109, no. 11, 2012, pp. 4169–4174. JSTOR, www.jstor.org/stable/41507114.
- Longo, Frank J., and Everett Anderson. “The Fine Structure of Pronuclear Development and Fusion in the Sea Urchin, Arbacia Punctulata.” The Journal of Cell Biology, vol. 39, no. 2, 1968, pp. 339–368. JSTOR, www.jstor.org/stable/1605485.
- Panda, Dulal, et al. “Differential Effects of Vinblastine on Polymerization and Dynamics at Opposite Microtubule Ends.” Journal of Biological Chemistry, vol. 271, no. 47, 1996, pp. 29807–29812., doi:10.1074/jbc.271.47.29807.
- Hamilton, E P et al. “Pronuclear fusion failure: an alternate conjugational pathway in Tetrahymena thermophila, induced by vinblastine.” Genetics vol. 118,4 (1988): 627-36.
External links
| Look up pronucleus in Wiktionary, the free dictionary. |