Polistes gallicus
Polistes gallicus (also historically referred to as Polistes foederatus) is a fairly common species of paper wasp found in various parts of Europe, excluding England, Denmark, and Scandinavia, from warmer climates to cooler regions north of the Alps.[1] The distribution of P. gallicus also extends into northern regions of Africa, Israel, Iran, and even parts of China and Russia.[2] Nests of these social insects are created in these various conditions. The Polistes species use an oral secretion to construct their nests, which consist of a combination of saliva and chewed plant fibers. This structural mixture physically protects the nest from various harsh elements and from weathering over time.[3]
| Polistes gallicus | |
|---|---|
 | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Arthropoda |
| Class: | Insecta |
| Order: | Hymenoptera |
| Family: | Vespidae |
| Subfamily: | Polistinae |
| Tribe: | Polistini |
| Genus: | Polistes |
| Species: | P. gallicus |
| Binomial name | |
| Polistes gallicus Linnaeus, 1761 | |
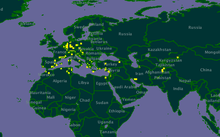 | |
| Distribution of P. gallicus in yellow | |
| Synonyms | |
| |

Description and identification
P. gallicus can be spotted due to its distinct markings of bright yellow and black. It is relatively small in size compared to other native Polistes species. Curled antennae are a common characteristic in the genus for males, despite being shorter in length in comparison to wasps of other species. Their faces are also completely yellow in color.[2] This species of wasp feeds its brood after visiting numerous flowers, collecting nectar in addition to feeding them meat. Due to its dimensions, they likely transfer pollen to the stigma, despite the fact that P. gallicus bodies are almost bald, which leads to few or no pollen grains stuck on them after foraging. This specific body type can help with identification.[4]
Taxonomy and phylogeny

P. gallicus is a member of the family Vespidae, further classified under the Polistinae (the second-largest of the subfamilies), which consists of various social wasps. Within the larger subfamily Polistnae, Polistes species are categorized by their independent founding behavior, distinguishing them from swarm-founding species.[5] P. gallicus is almost indistinguishable from the European paper wasp, P. dominula, with which it had long been confused. Nearly every reference to P. gallicus prior to 1985 was actually referring to P. dominula; due to the great similarity between species, however, many published studies for which no vouchered reference specimens are available cannot be reliably assigned to either species. Most of the literature that pertains to actual P. gallicus uses the junior synonym, P. foederatus, a name which still occasionally appears in the literature despite the known error.[1] It is also closely related to P. biglumis and P. hellenicus.[6]
Distribution and habitat
The extensive range of P. gallicus includes a variety of climates and habitats, although ideally it prefers to nest in warm and dry areas. In Italy, nests are typically built in open areas hanging from branches with the cells opened towards the ground. In the colder conditions north of the Alps, metal scraps such as pipes serve as protective enclosures to P. gallicus nests. These wasps are the most widely distributed Polistes in Spain. P. gallicus also inhabits parts of Paris, although the farther north, the more rare this species becomes.[2] Its conservation status in general has not been evaluated yet, although in its inhabited regions, it is common.[7]
Nest structure can consist of numerous cells, with some nests containing as many as 500 cells. These nests are constructed from a mix of oral secretions and plant fibers to make a paper pulp. Bigger nests are more commonly found in southern regions. For example, a study of two wasp nests in Cambridge, Massachusetts, found a nest constructed inside a stop sign pole that was 8 cm high and 5 cm wide and consisted of 134 cells. A second nest, that contained 153 cells within the same nest dimensions, was found suspended from a pipe.[2] The nests of P. gallicus tend to be smaller than those of comparative Polistes. Nest size also depends on the location, with nests in more sheltered areas tending to be larger.[3]
Colony cycle

In the spring, foundresses come out of hibernation (mid may) and form nests on their own[8] (late May to early April). Sometimes, two or more foundresses cooperate to a single nest, and nests with multiple founders can be found in southern regions of Germany and in Italy. But most P. gallicus colonies are initiated by a single foundress. The cells formed are usually in a hexagonal pattern, and are attached to a support by a single stalk. The material is collected from sticks and bush branches. The foundress then proceeds to lay eggs directly in the brood cells. Each cell will host a single egg, that takes 2 weeks to hatch. The newly hatched larvae are fed throughout their development, growing through 5 instars and then pupating. The mature larva (2 weeks old) spins a cocoon with which it encloses itself in the cell, and then proceeds to complete metamorphosis (other 10 to 14 days). Each of the season's first brood of wasps is exclusively females. These new members of the nest are workers and serve as subordinates to the foundress.[9]
In about 40 days, (May to early June) the first workers emerge, attending to tasks such as maintaining the nest and taking care of the brood inside, or defending it if threatened. The first workers take over all the duties in the colony, hunting for preys (larvae are fed with other insects that adults hunt) and collecting water, carbohydrates and nest material. The foundress is now a queen, that lays eggs and rests all day long while her workers do all the other work. More workers emerge, and the number of wasps soon increases. Newly hatched workers are bigger than the first ones, because they were well fed as larvae (unlike the first generation, reared solely by a single hard working wasp queen). In June and July, the colony grows and new cells are built. The nest can reach an extraordinary size for this genus, containing up to some thousand cells and a hundred of workers. But more often, this size is never reached. The time of development of brood contracts. The eggs laid from June on develop throughout a single month due to the warmer summer conditions and the increased availability of food, provided by an ever-increasing amount of restless workers. Among those eggs, there are several unfertilized ones that the queen willingly chooses not to fertilize. Those eggs will mature in males, while the others develop in larger females that don't cooperate to the nest. Those are the future foundresses, that rapidly develop their ovaries and the amount of fat in their body. The reproductive phase happens when those females emerge, in July to early August. Males generally emerge first, because their development requires a minor amount of time and food. Soon after August, the month of reproductive wasps rearing, worker numbers start dropping due to a decline in brood care as the older individuals start dying and replacements no longer exist. The initial foundress dies, while some workers start developing ovaries and laying eggs themselves. They are unfertilized, so their offspring will exclusively result in other males, if anything. The new foundresses and males gather in separate and unisexual clusters, and occasionally mate with non-relatives that they meet while feeding on flowers. Those clusters are common until November, often not too far from the nest, that no longer produces new wasps. All the workers die out, then the males, and only the fertilized foundresses overwinter in various shelters until the next year, when they start new colonies and the cycle goes on. The amount of foundresses that a mature colony can produce is high, up to a pair of dozens or more, in order for the species to cope with to threats and colony disturbance. A major threat to colonies in the pre-emergence phase (prior to the first workers) is ants. Those of the general "Tapinoma", "Pheidole" or "Tetramorium" can completely destroy a young nest in order to feed on larvae and pupae, and the foundress has to found a new nest from scratches or steal a nest from another foundress. Nest predation and queen usurpation in this species are very common, as well as premature death of the foundress/Queen. In this case, workers reproduce and rear their own offspring, made of males and doomed to extinction, without producing reproductive females. Nest refoundation by colonies that lose the original one is pretty common, and allows this species of wasps to thrive in highly disturbed landscapes, that face frequent fires or other disturbances. When the nest is destroyed or goes lost, the swarm just moves to a near point (a branch or else) and begins another nest. Sometimes, when there are lots of wasps, the swarm may break up and create several new little nests. Only the one with the foundress in it will produce female offspring and complete the cycle, while the other ones will end up producing worker males. If one of those secondary nests is lost, its workers will rather rejoin a near group of other sisters instead of trying to found it again.
Behavior
Diet

The wasp is omnivorous, feeding on fruits, flower's nectar, insects, snails, or larger animals' corpses.
Dominance hierarchy
Hormones play a role in the establishment of dominance hierarchies among P. gallicus. Dominant females tend to have more developed ovaries due to higher activity levels in their endocrine systems. A larger corpora allata also influences the determination of dominance. A higher reproductive capacity is indicative of the dominant female.[8]
Reproductive suppression
After a hierarchy is established, the dominant wasp remains the sole reproducer in the colony due to the inhibition of endocrine activity within the subordinate wasps. Various factors contribute to the possibility of inhibition, which might also lead to differences in endocrine activity. If subordinates happen to lay eggs after the hierarchy is formed, the dominant foundress will eat the eggs to ensure all laid eggs are of her own genes.[8]
Kin selection
.jpg)
Genetic relatedness within colonies
Colonies of P. gallicus have one queen that mates and produces offspring. All females, both workers and queens, are capable of producing male offspring, and have a genetic incentive to do so; mother-to-son relatedness is 1/2, while the relatedness of a worker to her brothers is 1/4. If the queen has only been mated once, then worker's relatedness to the male offspring of other workers is 3/8, meaning workers should favor male production by themselves and their sisters over production by the queen. However, in the case that the queen mates more than once, workers are actually more related to their brothers than compared to the offspring of other workers (a mixture of full and half sisters).[10]
Kin recognition and discrimination
P. gallicus reacts to chemical signals within Van der Vecht (VVS) organ secretions. VVS was studied and found to be a composition of hydrocarbons. The exact mixture tended to vary among differing colonies and also between the queen and workers, demonstrating worker specificity. Due to the different composition of the VVS chemicals, workers are able to distinguish these differences and respond with varying degrees of aggressiveness to other wasps which also release these chemicals. These chemicals can allow the wasps to interpret these cues about the presence of a related queen in the colony. While workers are able to discriminate VVS of their own queen and other workers, they are also able to distinguish odors when secreted from alien individuals.[11]
The foundress of the nest has unicellular glands associated with this organ that peaks in activity around the time when workers start emerging. These chemicals are also hypothesized to serve as repellent secretions to defend the nest before it is populated. Also beyond just serving as a means of communication, when the queen deposits VVS on the nest during a particular stroking behavior of the queen, this serves as an indication of ownership tied to the presence of this particular queen. It actually has an inhibitory effect on the ovarian development of other workers within the colony.[11]
Worker-queen conflict
Queen's interest
Due to the differences in relatedness within colonies, conflict over sex ratio arises. It would be beneficial for the queen to produce an equal ratio of sons and daughters due to equal relatedness between genders. Fisher's theory of equal investment supports this 50:50 sex ratio of males and females. This is a stable situation, as both males and females have the same expected reproductive success.[12] To maintain balance, queens may eat other workers' laid eggs if she was not successful in preventing workers from laying eggs in the first place.[10]
Workers' interest
For workers, the production of males by the queen would be more favorable than to let other workers produce progeny. This would lead to an increased relatedness around 3/4, compared to the relatedness of worker produced males which is around 2/5.[10] This might support the worker policing prediction that workers might prevent other workers from laying eggs in support of queen produced progeny when they are multiply mated.[13] In addition, in colonies with the presence of a queen, workers will not produce males, but instead sacrifice their own interests for the greater good of colony productivity. Indirect evidence supports matricide within the colony, and queen death has been noted to be high in P. gallicus.[10]
Interaction with other species
Parasites
Strepsipterans belonging to the genus Xenos are known to infect Polistes. Xenos vesparum has been especially documented in these wasps. Younger members of the colony are typically the target and most susceptible to parasitism. It is more difficult to document parasitized adult members due to their departure from the cell, which may play a role in perceived susceptibility. However, parasites are most visible in the neotenic adult or pupal stage. This does not distract from finding of strepsipteran parasites in all stages of the host, from egg, to larval, pupal, and finally the adult stage. Also, nests tended to have more parasites per brood member when parasite prevalence is high. Xenos species tend to infect these wasps either through phoretic transport in which the first-instar larvae are able to attach to wasps’ abdomens at flowering patches or by infecting masses by releasing close to combs from an infected wasp. Sometimes in brood with high levels of parasitized larval hosts, an adaptation of the parasite to enter the eggs might exist.[14]
Defense
Polistes species are known to use stings and venom as a means of colony defense. However, this venom seems to be costly to produce as they only release it after the sting in certain situations. Dangerous stimuli must first be perceived before they go out of their way, leaving a nest unattended, to attack.[15] In some situations, P. gallicus ise known to exhibit aggressive behavior to wasps of a foreign colony.[11] While venom from a basic standpoint is used by solitary species to capture prey, it has served a greater purpose of defense in social colonies against colony vertebrate and invertebrate offenders.[15]
When it comes to alarm systems, Polistes species can communicate with others through vibrational and visual signals. It might actually be beneficial for a smaller colony of wasps to switch from alarm pheromones, common with these wasps, to these alternative signals when the colony grows in size. Alarm pheromones mixed with the composition of the venom can also be released. However, if this release occurs following the act of ejecting venom by the signaling wasps or if it is due to the actual release of the venom during the sting has yet to be determined .[15]
Human importance
Gains from the study of Polistes venom
Knowing venom chemistry from these species of wasps can lead to human advantages for pharmaceuticals. The chemical breakdown of venom allows for synthesis in immunology therapy due to the creation of more reliable and effective treatments for people with allergies. Studies which analyzed the way venom interacts with victims provided a mechanism for drugs to permeate cell membranes. Further studies on wasps could provide a mechanism to control overpopulation through the creation of artificial sex attractants.[15]
Allergic reaction-induced IgE-mediated anaphylaxis is commonly a result of hymenopteran stings. The composition of venom from a sting can even affect the types of treatment a patient should be given. Differences have been found between the composition of American and European Polistes venoms. Response to different epitope spectrums depends on the type of Polistes that did the stinging. P. gallicus venom was found to be a combination of four major allergens: Ag5 (antigen 5), hyaluronidase, phospholipase, and protease. This discovery has led to the addition of these allergens into a standard Polistes mix containing venom from North American species to improve diagnosis and therapy for European patients with Polistes allergies.[16]
References
- O’Donnell, Sean (1998). "Reproductive caste determination in eusocial wasps (Hymenoptera: Vespidae )" (PDF). Annual Review of Entomology. 43 (1): 323–346. doi:10.1146/annurev.ento.43.1.323. PMID 15012393. Retrieved 23 September 2014.
- Hathaway, M.A. (1981). "Polistes gallicus in Massachusetts (Hymenoptera: Vespidae)". Psyche: A Journal of Entomology. 88 (1–2): 169–173. doi:10.1155/1981/94216. Retrieved 17 September 2014.
- Bagriacik, Nil (2012). "Comparison of the nest materials of Polistes Gallicus (L.), Polistes dominulus (Christ) and Polistes nimpha (Christ) (Hymenoptera: Vespidae)" (PDF). Arch. Biol. Sci. 64 (3): 1079–1084. doi:10.2298/abs1203079b. Retrieved 20 September 2014.
- Dafni, A.; R. Dukas (1986). "Insect and wind pollination in Urginea maritima (Liliaceae)". Plant Systematics and Evolution. 154 (1–2): 1–10. doi:10.1007/bf00984864.
- Arevalo, Elisabeth; Yong Zhu; James M Carpenter; Joan E Strassman (2004). "The phylogeny of the social wasp subfamily Polistinae: evidence from microsatellite flanking sequences, mitochondrial COI sequence, and morphological characters". BMC Evolutionary Biology. 4 (8): 8. doi:10.1186/1471-2148-4-8. PMC 385225. PMID 15070433.
- Larch, Rainer; Hannes Baur; Gaston-Denis Guex; Christophe Praz (2004). "A new species of the paper wasp genus Polistes (Hymenoptera, Vespidae, Polistinae) in Europe revealed by morphometrics and molecular analyses". ZooKeys (400): 67–118. doi:10.3897/zookeys.400.6611. PMC 4023243. PMID 24843256.
- "Polistes Gallicus (Linnaeus, 1767)." Polistes Gallicus. European Environment Agency, n.d. Web. 22 Sept. 2014. <http://eunis.eea.europa.eu/species/216123>
- Röseler, Peter-Frank; Ingeborg Röseler; Alain Strambi; Roger Augier (1984). "Influence of Insect Hormones on the Establishment of Dominance Hierarchies among Foundresses of the Paper Wasp, Polistes Gallicus". Behavioral Ecology and Sociobiology. 15 (2): 133–142. doi:10.1007/bf00299381. JSTOR 4599709.
- Hudson, K. Reeve (1991). "Polistes". In Kenneth G. Ross & Robert W. Mathew. The Social Biology of Wasps. Cornell University Press. pp. 99–148. ISBN 978-0-8014-9906-7.
- Strassmann, JE; Nguyen JS; Arévalo E; Cervo R; Zacchi F; et al. (2003). "Worker interest and male production in Polistes gallicus, a Mediterranean social wasp" (PDF). Journal of Evolutionary Biology. 16 (2): 254–259. doi:10.1046/j.1420-9101.2003.00516.x. PMID 14635864. Retrieved 18 September 2014.
- Dapporto, Leonardo; Antonio Santini; Francesca R. Dani; Stefano Turillazzi (2007). "Workers of a Polistes Paper Wasp Detect the Presence of Their Queen by Chemical Cues". Chemical Senses. 32 (8): 795–802. doi:10.1093/chemse/bjm047. PMID 17644826. Retrieved 23 September 2014.
- Davies, N. B., J. R. Krebs, and Stuart A. West. An Introduction to Behavioural Ecology. Oxford: Wiley-Blackwell, 2012. 978-1-4051-1416-5 pp367-389. Print.
- Ratnieks, F.L.W. (August 1988). "Reproductive harmony via mutual policing by workers in eusocial hymenoptera". The American Naturalist. 132 (2): 217–236. doi:10.1086/284846. JSTOR 2461867.
- Hughes, D.P.; J. Kathirithamby; L. Beani (2004). "Prevalence of the parasite Strepsiptera in adult Polistes wasps: field collections and literature overview". Ethology, Ecology and Evolution. 16 (4): 363–375. doi:10.1080/08927014.2004.9522627.
- Turillazzi, Stefano (December 2006). "Polistes venom: a multifunctional secretion" (PDF). Annales Zoologici Fennici. 43 (5–6): 488–499. Retrieved 24 September 2014.
- Pantera, Barbara; Donald R. Hoffman; Lara Carresi; Gianni Cappugi; Stefano Turillazzi; Giampaolo Manao; Maurizio Severino (2003-10-13). "Characterization of the major allergens purified from the venom of the paper wasp Polistes gallicus". Biochimica et Biophysica Acta (BBA) - General Subjects. 1623 (2): 72–81. doi:10.1016/j.bbagen.2003.07.001. PMID 14572904.